как разбудить и активировать спящие волосяные фолликулы на голове?
Густые, красивые, здоровые волосы — мечта, пожалуй, любой женщины. Но иногда они не просто выпадают активней, чем обычно, но и перестают восстанавливаться — на месте выпавшего не вырастает новый волос, поскольку волосяная луковица «засыпает». Отчего такое происходит и как «разбудить» волосяные фолликулы?
Почему «засыпают» волосяные фолликулы?
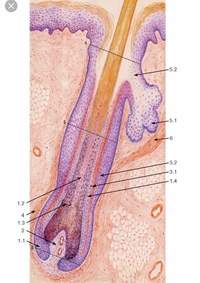
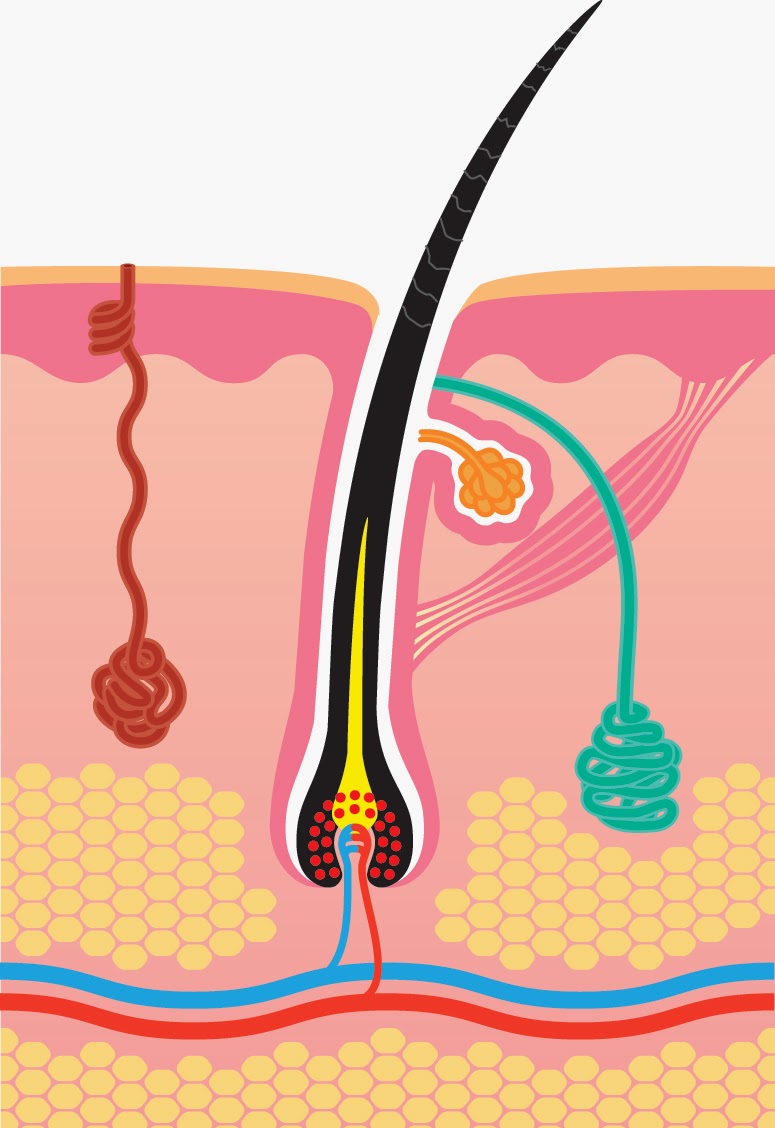
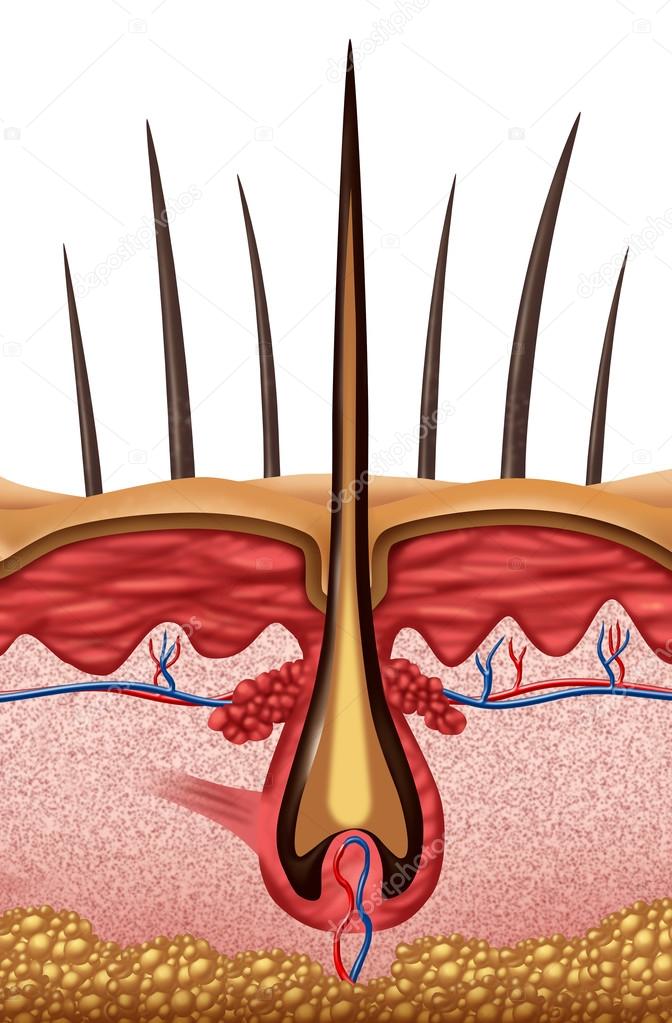
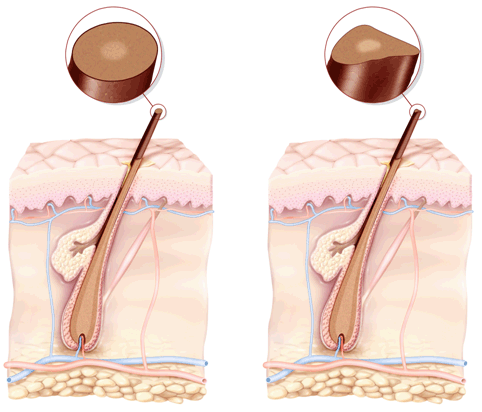
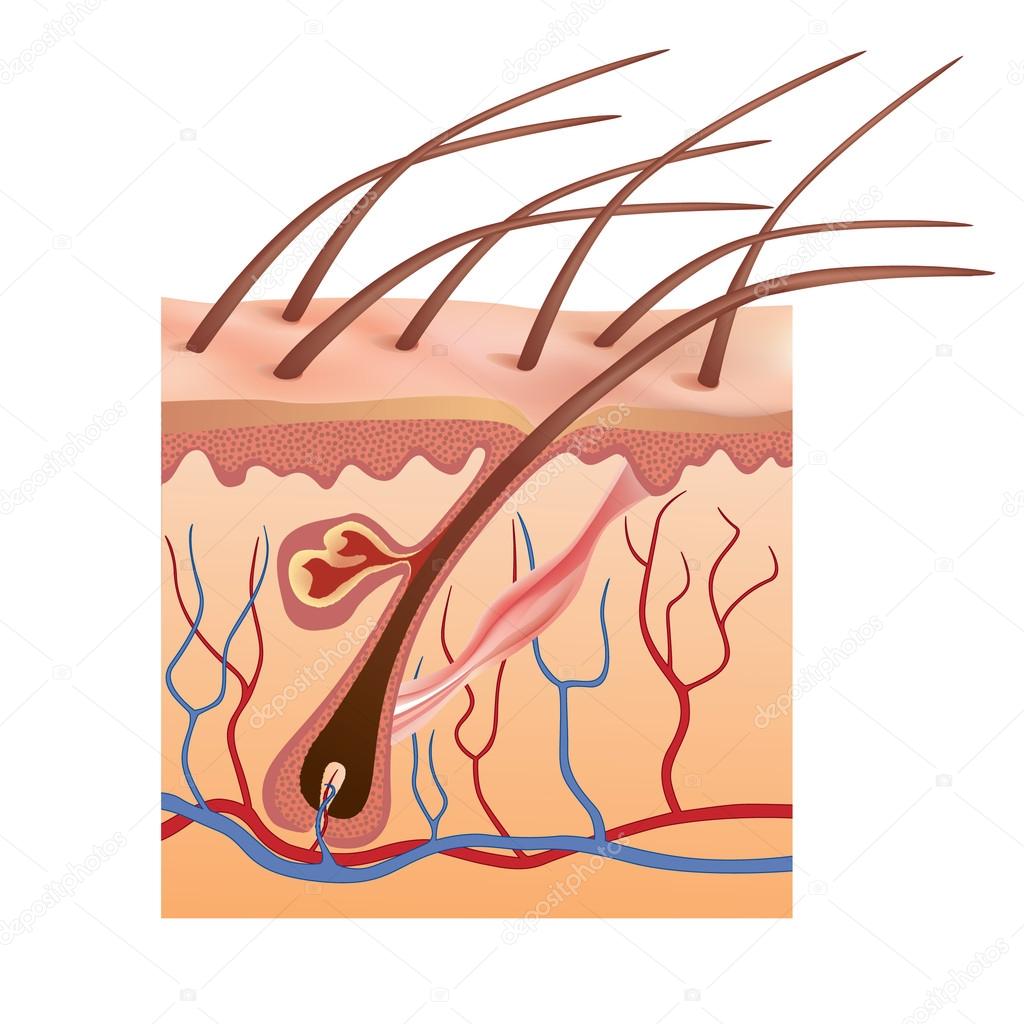
Волос состоит из стержня и луковицы. Стержень волоса — это та его часть, которая, собственно, и создает шевелюру. Но чтобы он появился, особые структуры внутри кожи должны выработать белки, из которых состоят волосы. Эти структуры, содержащие сложный комплекс элементов, называются волосяной луковицей. В ее состав входят:
- волосяной фолликул — комплекс клеток, вырабатывающий белок кератин, который составляет основу волоса;
- сальная железа — создает естественную среду, которая защищает кожу головы и волосы от потери влаги и микробов;
- сосуды и нервы, регулирующие питание волосяного фолликула.

В цикле жизни любого волоса есть несколько фаз.
- Анагенез, или активный рост. В ранней стадии этой фазы в волосяной луковице формируется молодой фолликул. Он начинает производить новый волос, и тот, отрастая, выталкивает старый (если он не выпал до того под действием внешних факторов). Фаза активного деления волоса продолжается в норме несколько лет — именно от ее длительности зависит, какой длины отрастают волосы, если их не стричь.
- Со временем активность деления сходит на нет, и наступает фаза катагенеза. Фолликул сокращается и начинает продвигаться к поверхности кожи. Но волосяной сосочек, из которого позднее сформируется новая волосяная луковица со всеми компонентами, остается на месте в спящем состоянии.
- Далее наступает фаза телогена, когда волос не растет и остается в дерме только потому, что его основание находится относительно глубоко. Именно такие волосы выпадают при расчесывании, мытье головы и других механических воздействиях. Эта фаза продолжается два–три месяца, после чего в нормальном состоянии волосяной сосочек «просыпается» и начинает формировать новую волосяную луковицу и волосяной фолликул.
 Эта фаза продолжается два–три месяца, после чего в нормальном состоянии волосяной сосочек «просыпается» и начинает формировать новую волосяную луковицу и волосяной фолликул.
Эта фаза продолжается два–три месяца, после чего в нормальном состоянии волосяной сосочек «просыпается» и начинает формировать новую волосяную луковицу и волосяной фолликул.Но иногда этого не происходит, и волосы остаются в спящем состоянии. Причин тому может быть немало:
- неправильное питание: недостаток белка, витаминов и минералов в пище. Если в организме начинает чего-то не хватать, он перенаправляет питательные вещества туда, где они необходимы прежде всего — например, на кроветворение. Волосяным фолликулам перестает хватать питания для нормальной жизнедеятельности, и они засыпают;
- болезни: острые и хронические инфекции, травмы, операции. Все это требует повышенного расхода питательных веществ и витаминов для восстановления организма. Волосы опять же начинают снабжаться по остаточному принципу;
- стрессы мало того что требуют повышенного питания для адаптации, так еще и перестраивают гормональный баланс, заставляя организм разрушать себя;
- гормональные изменения. Недостаточная функция щитовидной железы часто вызывает повышенное выпадение волос и переход волосяных луковиц в спящее состояние;
- воспаление кожи головы. Самая частая причина воспаления — себорейный дерматит, когда на коже размножаются особые грибки, вызывающие перхоть. Такое состояние нужно лечить специальными шампунями, содержащими противогрибковые средства. После того как воспаление стихнет, рост волос восстановится.
 Недостаточная функция щитовидной железы часто вызывает повышенное выпадение волос и переход волосяных луковиц в спящее состояние;
Недостаточная функция щитовидной железы часто вызывает повышенное выпадение волос и переход волосяных луковиц в спящее состояние;Вопреки обыденным представлениям, краска для волос, сушка феном, агрессивные средства для укладки не могут повредить волосяные луковицы или заставить «спать» волосяные фолликулы. Те находятся слишком глубоко и практически не подвергаются внешним воздействиям. Настолько глубоко, что практически нереально сжечь кожу краской для волос или обжечь горячим воздухом из фена. В этих случаях прическа редеет, поскольку волосы теряют прочность и эластичность и начинают обламываться зачастую довольно близко к коже. Но если прекратить агрессивно воздействовать на волосы и дать им время восстановиться и отрасти, становится заметно, что густота волос в принципе не изменилась.
Как «разбудить» волосяные луковицы: домашние рецепты
Витаминно-минеральные комплексы помогут восполнить дефицит витаминов и минералов в организме. Все эти вещества так или иначе взаимодействуют друг с другом, усиливая или, наоборот, взаимно ослабляя воздействие. Поэтому лучше не подбирать витамины самостоятельно, а предпочесть витаминно-минеральный комплекс, созданный с учетом этих взаимодействий. К тому же при различных изменениях в организме усиливается потребность в разных витаминах и минералах. Так что лучше не пытаться угадать самостоятельно, в чем же проблема, а выбрать готовый витаминно-минеральный комплекс для роста волос и ногтей. Хорошо, если комплекс содержит кератин или биотин (к примеру, Пантовигар®, «КОМПЛИВИТ® Формула роста волос»), защищающие волосы от воздействий внешней среды и придающие им упругость и блеск. Также есть БАДы, содержащие инозит, способствующий пробуждению волосяных фолликулов, и экстракт карликовой пальмы («Компливит® Формула роста волос»).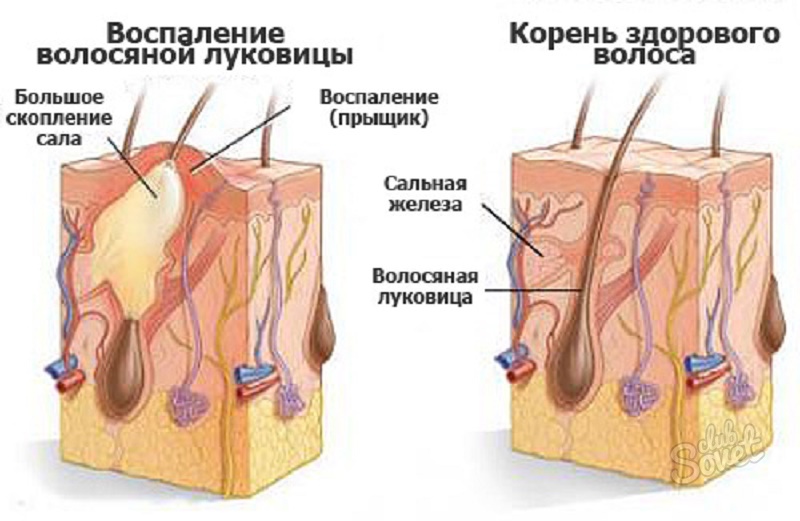
Диета. Для волос, как и для кожи, костей или мышц, нужны «строительные материалы», прежде всего полноценные белки. Поэтому правильное питание крайне важно для пробуждения волосяных луковиц. Постное мясо и нежирная птица, молочные продукты — главные источники белка для нашего организма. Строгим вегетарианцам можно обратить внимание на соевый белок и продукты из него.
Массаж, в том числе и самомассаж, — отличный способ улучшить кровоток в коже головы, а значит, увеличить доставку питательных веществ к волосяной луковице.
Никакие маски, шампуни, бальзамы, сыворотки и подобные продукты неспособны повлиять на волосяные фолликулы: те находятся слишком глубоко в коже, за естественным барьером, защищающим от проникновения любых веществ. Перцовые настойки, горчица и подобные средства хоть и усиливают кровоток, но сильно раздражают кожу головы, нарушают нормальный водно-липидный баланс и баланс микрофлоры, поэтому польза от них тоже сомнительна.
Активация спящих фолликулов с помощью косметических процедур
Пробудить спящие волосяные луковицы могут и некоторые салонные процедуры.
Прежде всего это внутрикожное введение лекарственных препаратов, например биоревитализация и мезотерапия, при которых в кожу головы тонкой иглой делаются инъекции специально подобранного комплекса витаминов и веществ, активирующих рост волос. Такие «коктейли» могут либо быть готовыми, производства известных косметологических компаний, либо создаваться непосредственно косметологом из нескольких лекарственных средств, набранных в один шприц. Плазмотерапия (PRP) — еще одна инъекционная методика, предусматривающая доставку живых тромбоцитов глубоко в ткани кожи.
Дарсонвализация кожи головы. Импульсным током раздражаются нервные окончания, расширяются сосуды, улучшается микроциркуляция, и, как следствие, активизируются волосяные луковицы.
Итак, неправильное питание, повышенная нагрузка на организм из-за стресса или болезней — вот основные причины, по которым волосяные луковицы надолго остаются в спящем состоянии. Единственный способ их пробудить — снабдить в достаточном количестве белком, витаминами и минералами, необходимыми для роста волос. Нужно помнить, что кожа — не слизистая желудочно-кишечного тракта и практически неспособна поглощать и доставлять в кровь питательные вещества. Поэтому самые верные способы доставить их к волосяным фолликулам — инъекции либо витаминно-минеральные комплексы, принятые внутрь.
Волосяной фолликул. Строение и фазы роста волос — Lazer Beauty
От количества фолликулов зависит густота волос на коже. Когда возникает чрезмерная густота волосяного покрова, избавиться от нежелательных волос поможет лазерная эпиляция
От количества фолликулов зависит густота волос на коже. Когда возникает чрезмерная густота волосяного покрова, избавиться от нежелательных волос поможет лазерная эпиляция — распространённая и востребованная процедура в косметологии. Удаление волос с помощью лазерной эпиляции считается самым радикальным и безопасным методом решения проблемы нежелательных волос. Стоит отметить, что лазерная эпиляция безболезненна и надёжна. Суть процедуры заключается в том, что вещество меланин, содержащийся в фолликуле, подвергается нагреву лазером, стволовые клетки волосяного мешочка нежелательных волос разрушаются, и эти волосы перестают расти. Импульса лазерного луча хватает на мгновенный нагрев, он действует избирательно, чтобы повлиять на структуру волосяных луковиц, не задевая близлежащие ткани. Поэтому лазерной эпиляции могут подвергаться такие чувствительные участки кожи, как зона лица, подмышек и бикини.
Удаление волос с помощью лазерной эпиляции считается самым радикальным и безопасным методом решения проблемы нежелательных волос. Стоит отметить, что лазерная эпиляция безболезненна и надёжна. Суть процедуры заключается в том, что вещество меланин, содержащийся в фолликуле, подвергается нагреву лазером, стволовые клетки волосяного мешочка нежелательных волос разрушаются, и эти волосы перестают расти. Импульса лазерного луча хватает на мгновенный нагрев, он действует избирательно, чтобы повлиять на структуру волосяных луковиц, не задевая близлежащие ткани. Поэтому лазерной эпиляции могут подвергаться такие чувствительные участки кожи, как зона лица, подмышек и бикини.
Если рассмотреть строение волоса и стадии его роста, то можно лучше понять, как воздействует на структуру волоса лазерный луч при процедуре эпиляции.
Волосяной фолликул и его строение
Волос состоит из стержня, который находится над кожей, и фолликула — невидимой части корня волоса, располагающейся в дерме, под поверхностным слоем кожи эпидермисом.
Волосяная воронка
Стержень волоса является продолжением волосяного корня. Он растёт из волосяной воронки — впадинки на коже. Ниже этой воронки стержень переходит в фолликул, состоящий из волосяного корня и прилегающих к нему тканей.
Волосяной мешочек
Слово «фолликул» в переводе с латинского означает «мешочек». Это особое углубление, в котором находится корень волоса. Его ещё называют корневым влагалищем. Волосяной мешочек книзу расширяется и образует волосяную луковицу.
Лазерный луч при эпиляции воздействует на стволовые клетки корня волоса в волосяной луковице, разрушая их и устраняя причину роста волос.
Другие структуры фолликула
К волосяному мешочку прилегают сальные и потовые железы, а чуть ниже — мышца волоса. Она приводит в движение волос под влиянием холода или эмоционального состояния человека, например, стресса или страха.
В самом низу фолликула имеется волосяной сосочек, состоящий из капилляров, соединительной ткани и нервных окончаний. Он выполняет питательную функцию, благодаря которой волосяные клетки получают все необходимые для роста волоса вещества.
Он выполняет питательную функцию, благодаря которой волосяные клетки получают все необходимые для роста волоса вещества.
Лазерная эпиляция и стадии активности фолликула
Волосяной фолликул с момента возникновения производит примерно 24-27 волос. Он проходит цикличные стадии активности и состояния покоя. От этих стадий зависят развитие и отмирание волоса.
Анаген — стадия наибольшей активности фолликула. В период анагена в волосяном корне происходит деление клеток. В результате появления новых клеток волос растёт. Этот период активного роста волос на голове длится от двух до пяти лет. Волосы бровей и ресниц растут от четырёх до семи месяцев. Около 85% волос находится в состоянии активного роста.
Катаген – это стадия, во время которой жизнедеятельность фолликула постепенно замирает. Волосяная луковица в это время отторгается от волосяного сосочка.
Телоген – период, когда в волосяном корне прекращается деление клеток. Старый волос с отмершей волосяной луковицей выпадает.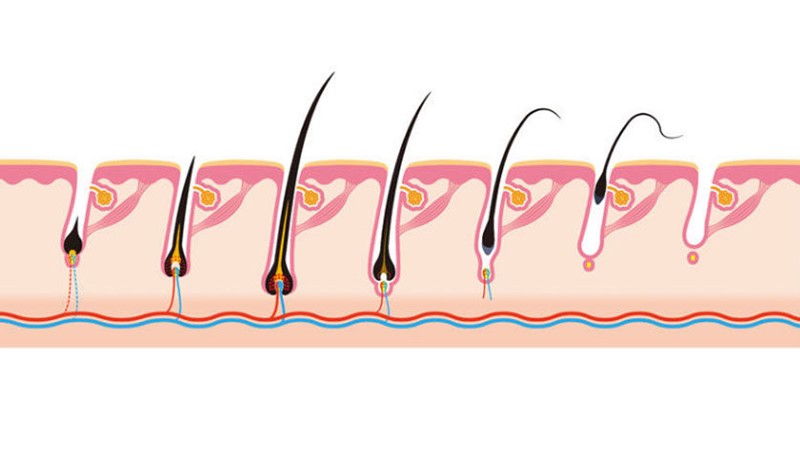
Затем стадия телогена постепенно переходит к анагену, стадии активности.
Лазерная эпиляция воздействует на нежелательные волосы исключительно на стадии анагена, активного цикла развития волосяного корня, когда в фолликуле происходит деление и образование новых клеток. Поэтому для большей эффективности процедура по удалению волос лазером должна проводиться несколько раз, так как волосы растут неравномерно.
Зачастую количество волос на разных стадиях активности и покоя зависит от густоты и состояния волосяного покрова, а также возраста пациента. Для получения максимального эффекта лучше всего чётко придерживаться советов и графика повторных процедур, назначенных опытным специалистом.
Лазерная эпиляция даёт мгновенный и долговременный результат, проблема роста нежелательных волос устраняется, а кожа становится гладкой и бархатистой.
Лечение волос, лечение выпадения волос, лечения облысения
ЛЕЧЕНИЕ ВОЛОС
Лечение алопеции (выпадение волос)
Прежде чем говорить о лечении алопеции (выпадения волос и облысения), предлагаем рассмотреть некоторые физиологические особенности роста волос.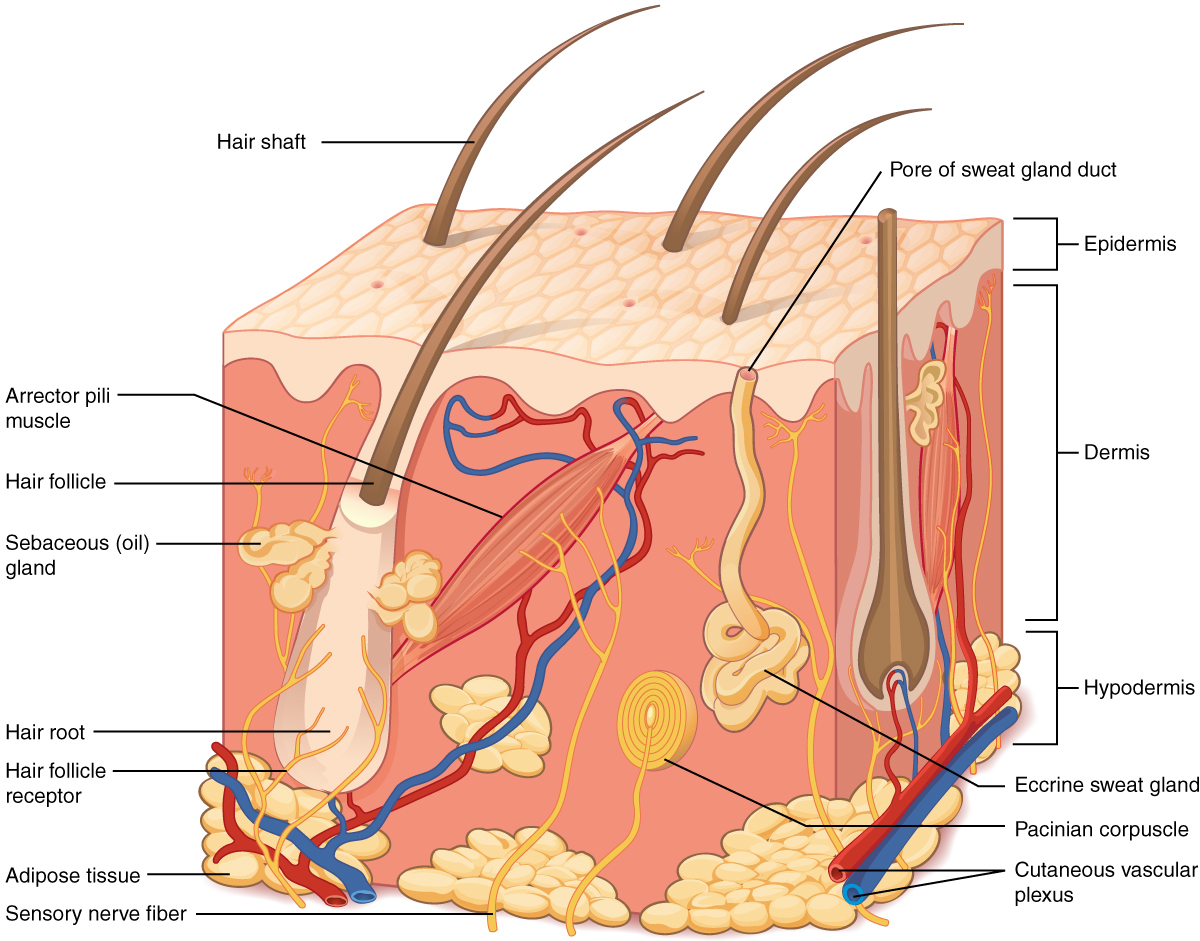
Волосяной фолликул функционирует циклично, циклы повторяются друг за другом:
- Анаген (фаза активного роста, т.е. жизни волоса)
- Катаген (переходная фаза)
- Телоген (фаза покоя, т.е.смерти волоса)
После того, как волос перешел в фазу телогена — он мертв, и единственный вариант развития событий — это выпадение этого телогенового волоса. На его месте развивается новый волос и далее цикл повторяется.
Луковица волоса является неотъемлемой частью самого волоса, соответственно, мертвый волос выпадает вместе с мертвой луковицей. Ее можно наблюдать в виде небольшого белесоватого утолщения на конце волоса. Это часто вызывает панику, т.к. многие считают это безвозвратным. Это не так. На месте выпавшего волоса формируется новый живой (анагеновый) волос с новой живой луковицей.
Волосяные фолликулы (ВФ) работают асинхронно (не одновременно). Это обеспечивает равномерную смену волос. Ежедневно часть волосяных фолликулов переходит в телоген (умирает), и примерно такое-же количество начинает новый цикл и входит в анаген. Фаза телогена длится 2.5 — 3.5. месяца. Если волос умер — он в течение этого времени будет находиться на голове, постепенно продвигаясь к поверхности кожи, и через 2.5 — 3.5 месяца выпадет.
Фаза телогена длится 2.5 — 3.5. месяца. Если волос умер — он в течение этого времени будет находиться на голове, постепенно продвигаясь к поверхности кожи, и через 2.5 — 3.5 месяца выпадет.
Минимальный срок для лечения выпадения волос — 3 месяца. Если пользоваться каким — либо средством только 1 месяц, то мы не вправе требовать результата. Телогеновый волос обязан будет завершить свой путь длиной в 3 месяца. Да, бывает, что через 2 — 3 недели лечения выпадение прекращается, но это другой диагноз и про такие легкие случаи мы не говорим, такое выпадение часто прекращается самопроизвольно.
Если беспокоит уменьшение объема волос (поредение), то эффект лечения волос Вы увидите тоже не скоро. Средняя скорость роста волос 1 — 1,5 см в месяц. Волосы должны успеть отрасти, чтобы сложиться в прическу, а это еще минимум 6 мес.
Если говорить об андрогенетическом процессе (сокращенно АГА), то о сроках лечения говорить в принципе не корректно. Это процесс хронический, и временное (курсовое) применение препаратов логично только на начальном (интенсивном ) этапе. После интенсивного курса должна оставаться какая-либо поддерживающая терапия (больше подходит словосочетание «специфический уход»). В противном случае, если отменить все средства, проблема очень быстро вернется. На консультации мы всегда говорим: лечение АГА после интенсивного курса — этот специфический уход за волосами будет ежедневной дополнительной процедурой наравне с чисткой зубов, бритьем (у мужчин), нанесением крема на кожу лица (у женщин).
Это процесс хронический, и временное (курсовое) применение препаратов логично только на начальном (интенсивном ) этапе. После интенсивного курса должна оставаться какая-либо поддерживающая терапия (больше подходит словосочетание «специфический уход»). В противном случае, если отменить все средства, проблема очень быстро вернется. На консультации мы всегда говорим: лечение АГА после интенсивного курса — этот специфический уход за волосами будет ежедневной дополнительной процедурой наравне с чисткой зубов, бритьем (у мужчин), нанесением крема на кожу лица (у женщин).
ЛЕЧЕНИЕ ГНЕЗДНОЙ АЛОПЕЦИИ. У пациентки в руках фотография до начала лечения.
Волосяные фолликулы • Ира Демина • Научная картинка дня на «Элементах» • Биология
На фото — волосяные фолликулы живой мыши (длина масштабного отрезка — 50 мкм). Зеленым флуоресцентным белком окрашены ядра эпителиальных клеток, красным — мезенхимальные клетки, в том числе волосяные, или дермальные, сосочки (в нижней части каждого из трех фолликулов). Фолликулы находятся в состоянии покоя, но готовы приступить к формированию нового волоса.
Фолликулы находятся в состоянии покоя, но готовы приступить к формированию нового волоса.
Волосяной сосочек (dermal papilla) богат кровеносными сосудами, за счет которых осуществляется питание клеток растущего волосяного фолликула, а также регенерация волоса — образование нового фолликула вместо утраченного. Если сосочек погиб, то волос больше не вырастет. Вокруг волосяного сосочка размещаются меланоциты — эпидермальные клетки, синтезирующие пигмент меланин, отвечающий за цвет волоса. Еще одна важная часть фолликула — так называемый бугорок (bulge) — участок между устьем сальной железы и местом прикрепления мышцы, поднимающей волос. В области бугорка находятся стволовые клетки.
Волосяные фолликулы — удобная модель для изучения стволовых клеток и их роли в процессе регенерации. В одном фолликуле за время его жизни последовательно формируется множество волос, при этом каждый раз фолликул проходит через процесс восстановления своей структуры после регрессии.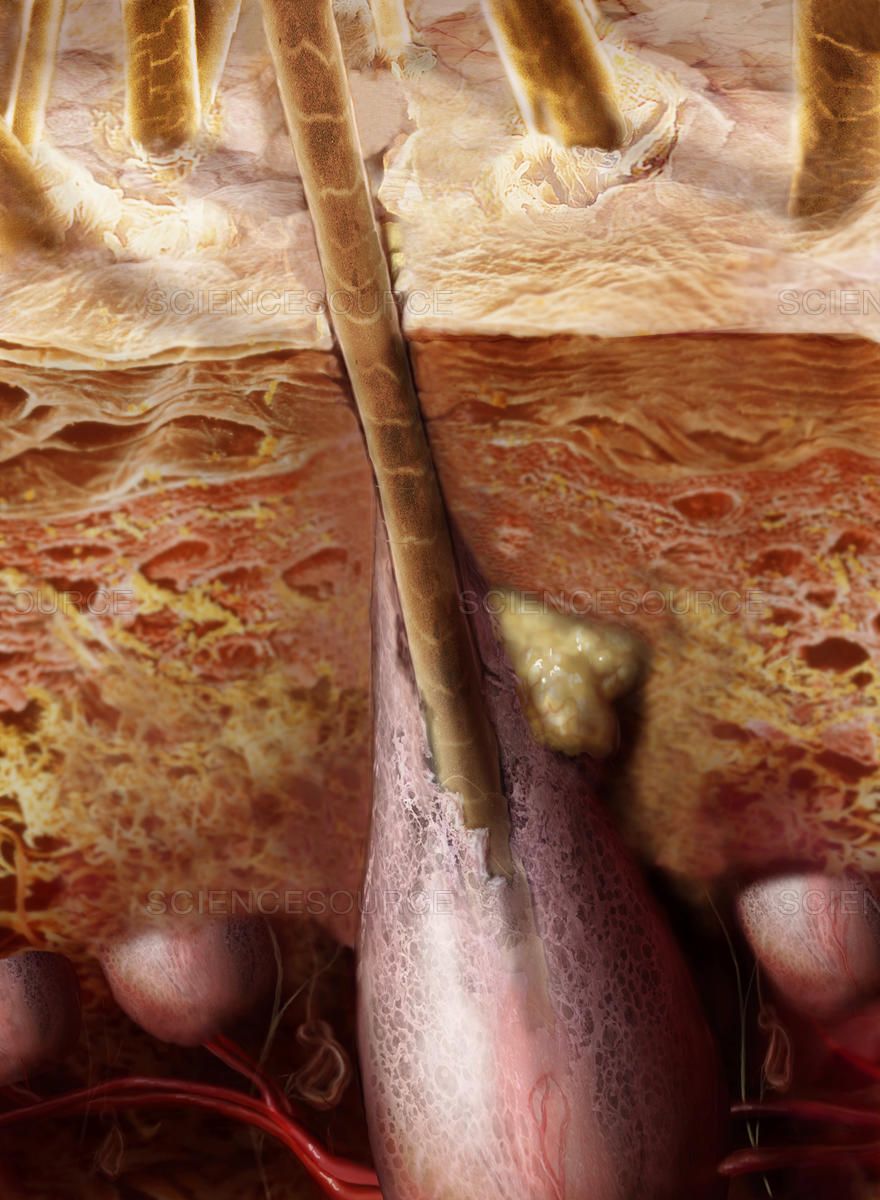 Методика прижизненной микроскопии позволяет отслеживать перемещение и пролиферацию разных типов клеток (в первую очередь, стволовых) в процессе регенерации фолликулов. Эти исследования важны для исследования поведения стволовых клеток при регенерации тканей, а кроме того, они помогают в разработке методов трансплантации волос при их утрате.
Методика прижизненной микроскопии позволяет отслеживать перемещение и пролиферацию разных типов клеток (в первую очередь, стволовых) в процессе регенерации фолликулов. Эти исследования важны для исследования поведения стволовых клеток при регенерации тканей, а кроме того, они помогают в разработке методов трансплантации волос при их утрате.
Для наблюдения за поведением клеток идеально подходят волосяные фолликулы на тыльной стороне уха мышей. Кожа здесь достаточно тонкая, фолликулы расположены неглубоко и под большим углом к поверхности кожи, что делает их удобными для микроскопирования. Кроме того, фолликулы здесь расположены достаточно плотно и имеют циклы развития с достаточно коротким периодом покоя перед фазой регенерации.
Для получения изображений оптических срезов кожи используют двухфотонный лазерный сканирующий микроскоп (TPLSM, two-photon laser scanning microscope). Благодаря неинвазивной технике микроскопирования можно наблюдать отдельные волосяные фолликулы в режиме реального времени.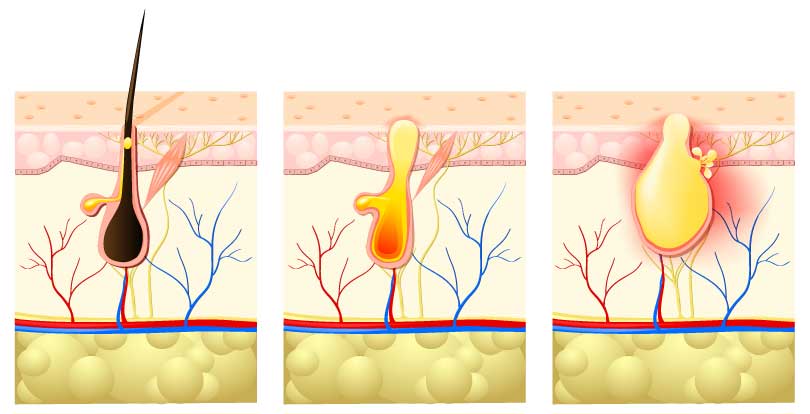 Ухо животного удобно каждый раз одинаково располагать на предметном столике TPLSM. При этом нужный участок кожи исследователи находят, ориентируясь на рисунок сосудистой сети на ухе и на точечные татуировки, которые делают на коже.
Ухо животного удобно каждый раз одинаково располагать на предметном столике TPLSM. При этом нужный участок кожи исследователи находят, ориентируясь на рисунок сосудистой сети на ухе и на точечные татуировки, которые делают на коже.
Волосяной фолликул в своем развитии последовательно проходит несколько стадий: анаген, катаген и телоген. Анаген — стадия активного роста волосяного фолликула, когда за счет интенсивного деления стволовых клеток формируется и растет стержень волоса. После анагена наступает катаген — стадия регрессии фолликула. Это промежуточный этап, во время которого рост волосяного стержня прекращается, на его нижнем конце формируется округлое уплотнение, а клетки фолликула между волосяным стержнем и дермальным сосочком отмирают. При этом волос лишается притока крови от дермального сосочка и полностью ороговевает, после чего фолликул переходит в стадию покоя (телоген). Ороговевший волос постепенно смещается вверх, к поверхности кожи и со временем выпадает, а в волосяном фолликуле после периода покоя начинается регенерация и образование нового волоса.
При механическом удалении волоса фолликул сразу переходит из катагена в анаген, поэтому когда исследователи сбривали волосы на ухе мыши (с дополнительным нанесением депиляционного крема), это стимулировало начало регенерации в покоящихся фолликулах. Продолжительность той или иной стадии развития фолликула зависит от вида животного, а также от местоположения фолликула на теле. Например, стадия телогена волосяных фолликулов головы человека длится около 3 месяцев, фолликулов бровей — 8–9 месяцев, а фолликулов ушей мыши — 2–3 недели.
Регенерация волосяного фолликула происходит за счет «резерва» стволовых клеток, которые сохраняются в фолликуле на протяжении всех стадий его развития. Как мы уже упоминали, стволовые клетки находятся в области бугорка. К концу анагена часть клеток бугорка перемещается ближе к основанию фолликула и начинает интенсивно делиться, оставаясь недифференцированными. Во время катагена группа таких клеток — так называемые клетки-потомки стволовых клеток бугорка — располагается в нижней части волосяного фолликула на границе с дермальным сосочком и образует «зародыш волоса» (hair germ). Эти клетки сохраняются во время телогена, и именно за счет них происходит дальнейшая регенерация волосяного фолликула и образование нового волоса в следующем цикле.
Эти клетки сохраняются во время телогена, и именно за счет них происходит дальнейшая регенерация волосяного фолликула и образование нового волоса в следующем цикле.
Исследования показали, что рост нового волоса инициируют клетки дермального сосочка. Они подают сигнал своим соседям — потомкам стволовых клеток бугорка, которые начинают усиленно делиться. Клетки сосочка способны вызывать рост волосяного фолликула de novo при трансплантации волос.
Благодаря неинвазивной технике микроскопирования удалось проследить в режиме реального времени развитие волосяных фолликулов с неповрежденным дермальным сосочком и без него. Исследователи проводили лазерную абляцию — разрушали клетки дермального сосочка с помощью лазера в самом начале стадии анагена, при этом стволовые клетки и их потомки оставались неповрежденными. В фолликулах с разрушенным мезенхимальным дермальным сосочком регенерация останавливается, что подтверждает необходимость этой структуры фолликула для его регенерации.
Видео разных этапов регенерации волосяных фолликулов мыши в режиме реального времени можно посмотреть здесь.
Фото из статьи C. Sedwick, 2014. Valentina Greco: Got hair? Длина масштабного отрезка — 50 мкм.
Ира Демина
Интересные факты о волосах
Шикарные волосы – это настоящее украшение, дарованное человеку природой. Но они не только дополняют наш образ, а еще и защищают кожу от холода и жары. Нет никакой растительности только на стопах, ладонях, губах и слизистых оболочках. Но что мы знаем о волосах, и все ли мнения являются истиной?
Неопровержимые факты
- Каждый волосок на теле имеет богатый минеральный состав: около 14 компонентов, присутствует золото, фосфор, железо.
- Здоровая копна в мокром виде очень эластична – растягивается до 30 % от первоначальной длины.
- Каждый человек теряет в день от 50 до 150 волосков.
- Существует 3 стадии роста, причем в активной стадии волос находится лишь 30-35% от всей своей «жизни», остальное время уходит на отмирание и выпадение.
- Здоровая копна, сплетенная в косу, способна выдерживать огромный вес – от 12 тонн (эквивалент 2 слонов)!
- Волос – настоящая «улика», которая способна многое рассказать о своем хозяине – тип питания, наличие наркотиков и алкоголя в организме, состояние крови, принадлежность к определенной расе.
- На теле человека столько же волос, сколько на теле шимпанзе – только наши волосы тоньше, короче и светлее.
Волнующие вопросы
В погоне за красивыми прическами женщины уверены, что могут сами повлиять на рост волос. К сожалению, это заблуждение. А в реальности:
- Частое подстригание кончиков волос никак не влияет на скорость их роста. Касается это и факта бритья наголо!
- У женщин волосы отрастают медленнее, чем у представителей сильного пола. Связано это с гормональным фоном.
- Ваша прическа будет становиться длиннее именно в теплое время года – волосяные фолликулы активизируются.
- Жизненный цикл волоса составляет от 2 до 7 лет.
- Волосяные фолликулы формируются у младенца еще в утробе матери.
Выпадение волос
Редеют волосы? В чем может быть причина:
- Неправильные щетки (особенно жесткие из металла) – лучше воспользуйтесь гребнем.
- Нарушение работы щитовидной железы – самая распространенная причина выпадения волос, но она обратима при правильной терапии.
- Нарушенный гормональный фон и неправильное скудное питание (диеты).
- При достижении 50 лет мужчины начинают активно терять волосы (около 50%), дамы столкнутся с проблемой в период менопаузы – 40%.
- Химическая завивка, частое воздействие высоких температур, ношение тугих резинок и заколок – нарушается кровообращение кожи головы, гибнут волосяные луковицы, соответственно и прическа может поредеть.
Наследственность
- Как ни парадоксально, но цвет и структура наших волос заложена генетически, и мы ничего не сможем изменить – все зависит от специальных красящих пигментов и генов, отвечающих за структуру будущей растительности на теле.

Все ближе к красоте!
Меры, которые позволят сохранить ваши волосы в отличном состоянии:
- Расчесывайте волосы часто – но только подходящей расческой и бережно.
- Чтобы волосы сияли здоровьем и красотой необходимо добавить в ежедневный рацион: зеленые овощи, витамины группы D и С, лосось, яйца, продукты, богатые фолиевой кислотой, йодом, морковь.
- Шампуни от перхоти способны сильно навредить волосам, они создают пленку (обволакивая волосок воском), сильно нарушая их структуру и предотвращая проникновения кислорода к луковице.
Цвет волос
- Обладателями светлых волос на планете являются только 2% людей. Это натуральные блондины.
- Брюнеты и шатены встречаются у людей чаще всего в мире.
- Рыжие – самый редкий оттенок, обладателями которого является только 1% всего населения на планете.
Забота о прическе
- Седые волосы могут появиться на голове после тяжелого стресса или в период старения буквально через 13 дней.
- Мужчины за всю жизнь тратят до 5 месяцев на бритье.
- Если не сбривать бороду никогда – длина достигнет примерно 9 метров.
Волосы растут всю жизнь, постоянно выпадают, погибают и процесс начинается заново, а вот волосяная луковица «умирает» раз и навсегда. Если ее уничтожить, на этом месте уже никогда не будет растительности. И если в отношении волос на голове такой ситуации нужно стараться всеми силами избегать, то вот исчезновение волос на теле – мечта многих. Благодаря лазерной эпиляции, которая, воздействуя на меланин, разрушает именно волосяные фолликулы, эта мечта может легко стать реальностью!
Дерматологи научились выращивать волосяные луковицы при помощи 3D-принтера
Первые операции по пересадке волос были проведены в 1939 году — пациентами дерматологов того времени стали солдаты Второй мировой войны, пострадавшие от сильных ожогов. С тех пор появилось как минимум два метода трансплантации волосяных луковиц из одной части тела в другую, но главная проблема технологии оставалась актуальной — операцию можно было проводить только тем людям, у которых есть откуда взять волосы. Теперь ситуация изменилась, потому что исследователям из Колумбийского университета впервые в истории удалось вырастить волосяные луковицы в лабораторных условиях.
Теперь ситуация изменилась, потому что исследователям из Колумбийского университета впервые в истории удалось вырастить волосяные луковицы в лабораторных условиях.
Новая технология имеет большие перспективы, и после коммерциализации сможет вернуть роскошную шевелюру миллионам мужчинам и женщинам из самых разных уголков мира. Количество донорских волосяных луковиц больше не будет играть никакой роли, потому что в лабораторных условиях их уже сейчас можно создать в бесконечном количестве. Более того, операции по пересадке волос в будущем могут вообще не пригодиться — имея большое количество образцов человеческих волос, производители средств от облысения смогут проводить больше экспериментов и разрабатывать более эффективные продукты.
Ученые вырастили искусственную волосяную луковицу
Новая технология выращивания волосяных луковиц состоит из нескольких частей. В самом начале исследователи методом 3D-печати создали объемную структуру из биосовместимого пластика — она послужила сосудом для роста волосяных луковиц и воссоздала подходящую для роста живых клеток микросреду.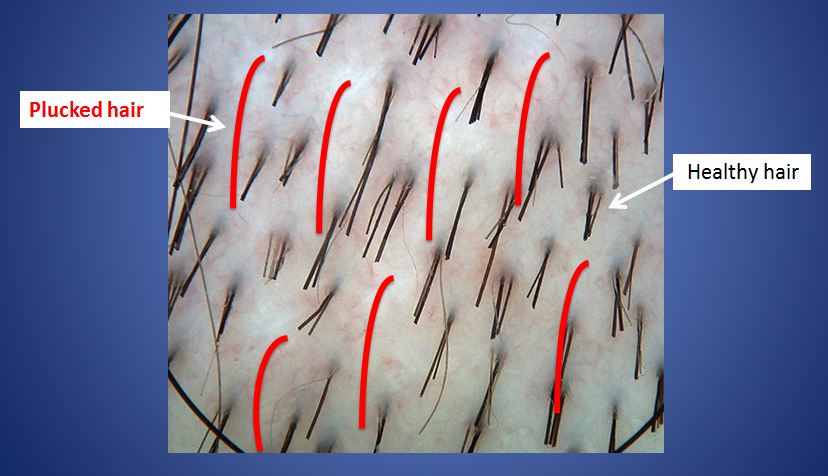 Каждый канал диаметром 0,5 миллиметра стимулировал образование луковиц, которые впоследствии помещались на выращенные клетки кожи.
Каждый канал диаметром 0,5 миллиметра стимулировал образование луковиц, которые впоследствии помещались на выращенные клетки кожи.
Эта своеобразная конструкция обрабатывалась сигнальными молекулами, которые подавили сигнальные пути под названием JAK-STAT. Из предыдущих исследований ученым известно, что они удерживают луковицы в неактивном состоянии, но благодаря молекулярной обработке они активизируются и волосяные луковицы начинают расти.
Исследователи считают, что при помощи технологии они могут создавать целые фермы волосяных луковиц. Полученные сетки с волосами впоследствии можно пересаживать на кожу головы, тем самым возвращая лысым людям волосяной покров. Результат такой трансплантации можно увидеть на видео ниже.
Порой от облысения спасают весьма неожиданные средства. Например, в 2016 году исследователи из Бразилии обнаружили, что для этого отлично подходит лекарства для лечения ревматоидного артрита.
Советуем почитать:
Пациентам с облысением вернули волосы с помощью препарата от ревматоидного артрита.
Обсудить эту, и другие темы науки и технологий, можно в нашем Telegram-чате. Также вы можете просто написать комментарий под материалом.
Можно ли стимулировать рост бороды?
Красота и здоровье
«Можно ли ускорить рост бороды?» – с таким вопросом Cook&Look обратился к экспертам школы парикмахерского искусства Simushka. «Конечно, можно!» – в один голос ответили они.
Вопрос в том, КАК вырастить бороду там, где она не растёт, и заставить её расти быстрее. Своими секретами в этом нелегком деле поделились:
Артём Апачек, основатель барбершопа Apaches Moscow, директор по барберингу в школе Simushka
Павел Брадобрей, энтузиаст и виртуоз опасного бритья, известен тем, что может безукоризненно побрить мужчину, например, топором или коньком, преподаватель Simushka
Елизавета Соколова, одна из первых барбер-girl России, владелец барбершопа Barberella, эксперт по влажному бритью, преподаватель Simushka
Секреты от Артёма Апачека:
Если борода растет, но в ней есть отдельные «пробелы»:
Помогут средства на основе масла красного перца, экстракта тропической ванили, богатые витаминами групп А и B и аминокислотами. Они разогревают кожу, стимулируя кровообращение. С кровью к коже поступают питательные микроэлементы, которые способны пробудить спящие волосяные луковицы. Важно наносить средства регулярно, от 1 до 3 месяцев массажными пощипывающими движениями против роста волос.
Они разогревают кожу, стимулируя кровообращение. С кровью к коже поступают питательные микроэлементы, которые способны пробудить спящие волосяные луковицы. Важно наносить средства регулярно, от 1 до 3 месяцев массажными пощипывающими движениями против роста волос.
Если в некоторых зонах борода не растет вообще:
У многих молодых людей борода на щеках не растет в принципе. Да, спящие волосяные луковицы там есть, но разбудить их бывает не под силу безопасным препаратам. Я, например, втирал средство для роста в течение нескольких месяцев и горжусь тем, что вместо 5 волос на левой щеке – стало 7. Хотя мой друг был настойчивее и втирал год – вот тогда начал появляться равномерный результат. Однако окладистой бороды после всех процедур ждать все равно не приходится, увы.
Если все же хочется бороду там, где ее нет:
Если вас очень беспокоит отсутствие растительности на лице, обратитесь к врачу-трихологу. Он даст вам верные рекомендации. В крайних случаях врачи назначают гормональные препараты на основе тестостерона.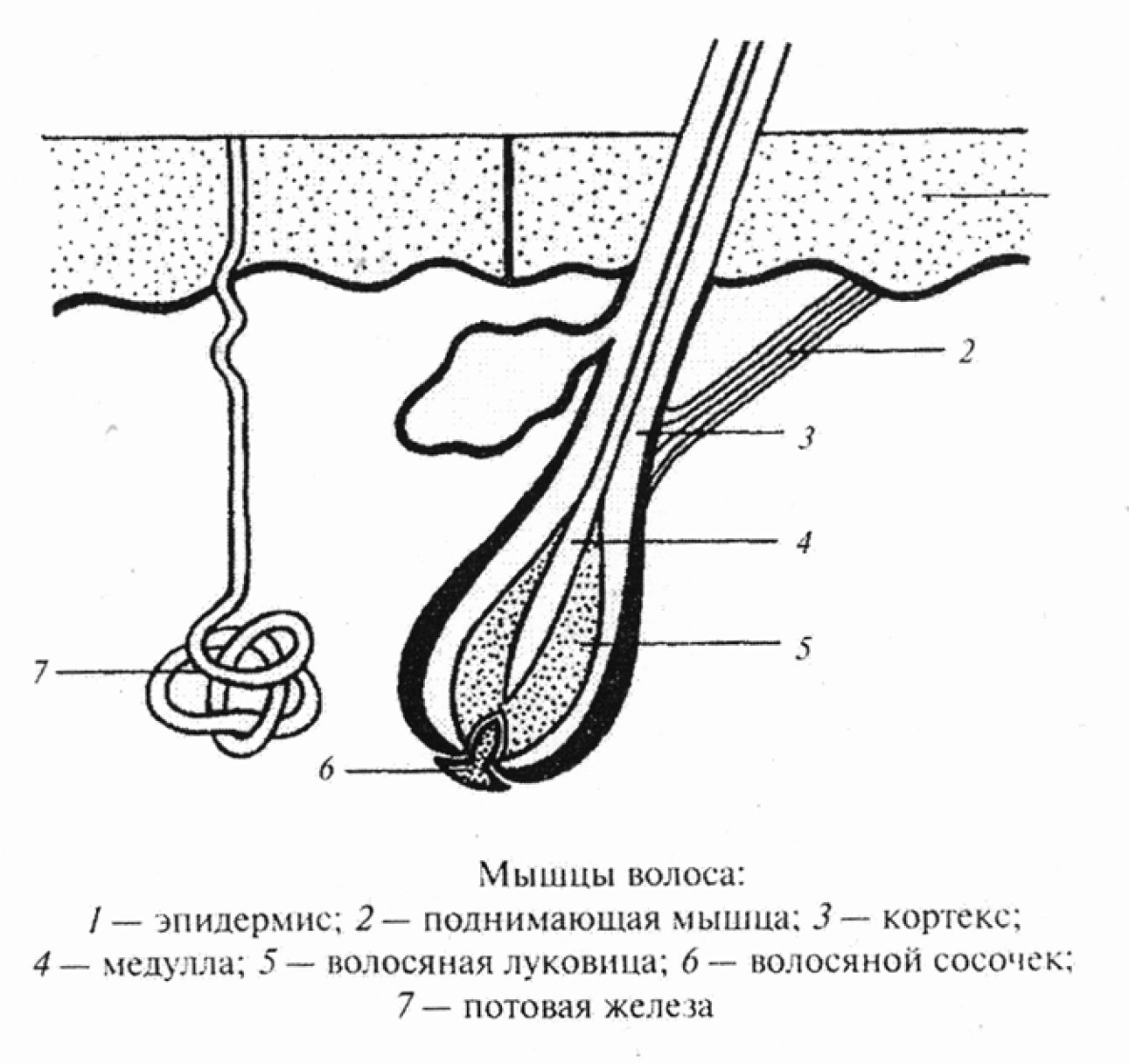 Ни в коем случае не начинайте их использование самостоятельно, это серьезная вещь, которая может негативно сказаться на вашем здоровье без контроля специалиста.
Ни в коем случае не начинайте их использование самостоятельно, это серьезная вещь, которая может негативно сказаться на вашем здоровье без контроля специалиста.
Секреты от Павла Брадобрея:
Для тех, кто хочет длинную бороду:
Чтобы борода росла быстрее, нужны питательные масла, например, масло виноградной косточки, ши, арганы. Нужно втирать их в кожу и распределять по длине бороды. И, конечно, нужны компрессы из горячих полотенец и добрые руки вашего барбера.
В домашних условиях Cook&Look рекомендует использовать электробритву – такую как Panasonic ES-SL41 с возможностью сухого и влажного бритья. Из дополнительных плюсов: три лезвия, заточка под углом 30 градусов, легко промывается водой и абсолютно безопасна.
Сеточная бритва Panasonic Cook&Look рекомендует использовать электробритву – такую как Panasonic ES-SL41 гарантирует отличный результат даже в борьбе с самой густой щетиной. Внутренние лезвия из японской стали Nanotech с заточкой 30 градусов, созданы с применением нанотехнологий и удаляют волосы у самых корней, обеспечивая максимальную гладкость кожи, не вызывая раздражения. Стильный корпус, сделанный на выбор в красном, серебристом и синем цветах, и невысокая стоимость бритвы делают ее доступной для всех любителей эффективного и качественного бритья!
Стильный корпус, сделанный на выбор в красном, серебристом и синем цветах, и невысокая стоимость бритвы делают ее доступной для всех любителей эффективного и качественного бритья!
Секреты от Елизаветы Соколовой:
Для тех, кто хочет бороду любой ценой:
В наше время существуют такие услуги, как пересадка волос на лицо и даже наращивание бороды.
Длинная здоровая борода требует системного подхода:
Для ускорения роста бороды действует тот же принцип, что и для ногтей и волос на голове – поможет только воздействие на организм в целом и изнутри. Это различные комплексы витаминов, которых на рынке сейчас большое количество. Существуют составы, специально предназначенные для этих целей. У меня есть клиенты, которые активно занимаются фитнесом и употребляют различные биодобавки – в первую очередь, для спортивных целей, но волосы растут быстрее – это как побочный эффект.
Полная структура, функции и связанные заболевания — SkinKraft
Ваши волосы — самая большая любовь вашей жизни, и вы не можете с ними расстаться, верно? Каждая прядь волос образована волосяными фолликулами, расположенными на внешнем слое кожи головы.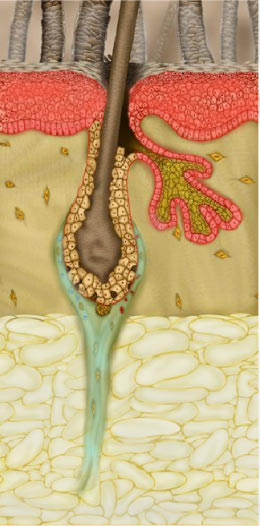 Это означает, что здоровье ваших волос напрямую связано со здоровьем кожи головы и волосяных фолликулов.
Это означает, что здоровье ваших волос напрямую связано со здоровьем кожи головы и волосяных фолликулов.
У среднего человека на коже 100 000 волосяных фолликулов. Таким образом, ваши волосяные фолликулы заслуживают особого внимания при уходе за волосами.Итак, продолжайте читать, чтобы понять структуру ваших волосяных фолликулов, их функции и проблемы с волосами.
Основные моменты:
Что такое волосяной фолликул?
Волосяные фолликулы [1] — это крошечные отверстия или поры в вашей коже. Их основная функция — отрастить волосы. На коже головы тоже есть волосяные фолликулы.
С биологической точки зрения волосяной фолликул выглядит как туннельная структура, расположенная в эпидермисе (внешнем слое кожи) [2]. Рост волос начинается у основания волосяного фолликула.
Корень ваших волос состоит из белка (кератина) [3] и питается кровью из кровеносных сосудов на коже. Кожный жир или натуральное масло, вырабатываемое сальными железами [4] возле волосяных фолликулов, сохраняет волосы и кожу головы смазанными.
Разрушитель мифов:
Теперь вы знаете, что волосы не растут из кончиков. Когда люди рекомендуют регулярную стрижку для получения более длинных волос, это ошибочное мнение. Регулярная стрижка может убрать секущиеся концы и повредить волосы, но не отрастит волосы.
Структура волосяного фолликула
Волосы состоят из двух частей — волосяного фолликула и стержня волоса. Волосяной фолликул фиксирует или удерживает волосы на коже головы. Он состоит из следующих частей [5]:
1. Лампа
Луковица находится у корня волос, где растут белковые клетки (кератин), из которых образуются волосы.
2. Папилла
Сосочек обеспечивает кровоснабжение волосяных фолликулов для здоровых волос.
3. Зародышевый матрикс
Зародышевый матрикс — это область, в которой клетки производят новые волоски.Он расположен в нижней части волосяного фолликула.
4. Выпуклость
Выпуклость расположена в середине волосяного фолликула. В нем есть стволовые клетки, которые регенерируют волосяные фолликулы, сальные железы и эпидермис. У выпуклости также есть arrector pili, мышечная ткань, из-за которой ваши волосы встают дыбом, когда вы испытываете мурашки по коже.
В нем есть стволовые клетки, которые регенерируют волосяные фолликулы, сальные железы и эпидермис. У выпуклости также есть arrector pili, мышечная ткань, из-за которой ваши волосы встают дыбом, когда вы испытываете мурашки по коже.
Функции волосяного фолликула
Помимо роста волос, волосяные фолликулы играют важную роль в принятии решения о том, как ваши волосы должны выглядеть.
Форма фолликула зависит от типа ваших волос.Круглые волосяные фолликулы делают волосы прямыми, а овальные — более кудрявыми.
Другая функция волосяного фолликула — придавать цвет вашим волосам. Волосы, как и ваша кожа, приобретают цвет за счет пигмента под названием меланин [6]. Этот меланин хранится в волосяных фолликулах. Есть два типа меланина — эумеланин и феомеланин.
- Если у вас много меланина, у вас будут черные волосы.
- Если эумеланин умеренный, ваши волосы коричневые.
- Если у вас очень мало меланина, ваши волосы светлые.
- Если у вас феомеланин, ваши волосы рыжие.
Способность волосяных фолликулов вырабатывать меланин снижается с возрастом, и в результате вы увидите седые или белые пряди волос.
Обычно волосы растут на полдюйма каждый месяц. На этот показатель влияет ваш
Обычно, если вы выдергиваете волосы из волосяного фолликула, он может вырастить новые волосы. При определенных заболеваниях волосяные фолликулы больше не могут расти и приводить к появлению лысин или пятен.
Цикл роста волос
Волосы из волосяного фолликула циклически растут на поверхность.
1. Анаген (фаза роста)
Это этап, который длится 3-7 лет. Волосы начинают расти от корня.
2. Категен (переходная фаза)
В этой фазе рост замедляется, и фолликул сокращается. Это может длиться до 2-4 месяцев.
3. Телоген (фаза покоя)
Старые волосы выпадают, и новые волосы снова начинают расти от корней.Эта фаза может длиться до 3-4 месяцев.
Не все фолликулы на коже головы находятся в одной и той же фазе в любой момент времени. Разные фолликулы проходят три разные стадии роста.
Разные фолликулы проходят три разные стадии роста.
У здорового человека 90% волосяных фолликулов находятся в фазе роста или анагена. Здоровый человек может терять около 100 прядей в день.
Болезни, связанные с волосяными фолликулами
1. Андрогенетическая алопеция
Андрогенетическая алопеция [7] или облысение по мужскому типу — это состояние, которое влияет на циклы роста волосяных фолликулов на коже головы.Цикл волос замедляется и останавливается. Фолликулы больше не могут производить волосы. Это также может повлиять на женщин и называется облысением по женскому типу, когда оно проявляется у женщин.
2. Alopecia Areata
Очаговая алопеция [8] — это аутоиммунное заболевание, означающее, что иммунная система организма атакует собственные волосяные фолликулы. В результате волосы выпадают клочьями. Стероидные инъекции и кремы для местного применения замедляют выпадение волос в этом состоянии, но нет лекарства, чтобы полностью остановить выпадение волос.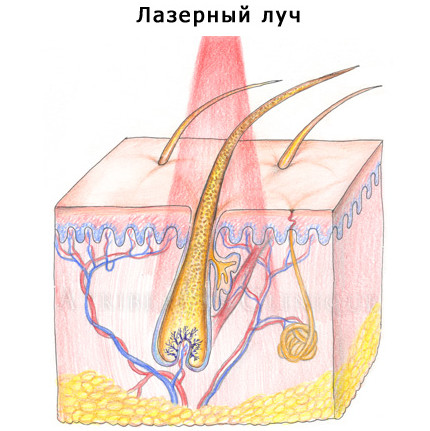 Алопеция иногда может привести к универсальной алопеции — полному выпадению волос по всему телу.
Алопеция иногда может привести к универсальной алопеции — полному выпадению волос по всему телу.
3. Фолликулит
Фолликулит [9] — это воспаление волосяных фолликулов. Фолликулит может поражать кожу головы, подмышки, лицо, руки или ноги. Фолликулит или инфекция волосяного фолликула проявляется в виде небольших красных, желтых или белых шишек или высыпаний и может содержать гной. Это вызвано стафилококковой инфекцией (бактериальной инфекцией).
4. Телоген эфлювий
Телогеновый отток [10] — временная форма выпадения волос.Посттравматический стресс может привести к преждевременному переходу фолликулов в фазу телогена или покоя. Роды, хирургическое вмешательство, болезнь, физическая или психическая травма могут вызвать отток телогена.
Примечание:
Проконсультируйтесь с дерматологом, если вы заметили внезапное и необычное выпадение волос или воспаление на коже головы.
Завершение
Волосяные фолликулы производят волосы в трех фазах роста. Они также определяют тип и цвет ваших волос.Поврежденные волосяные фолликулы могут вызвать воспаление или выпадение волос. Таким образом, понимание науки, лежащей в основе ваших волосяных фолликулов, и выбор правильного режима ухода за волосами имеет решающее значение, если вы хотите здоровые волосы.
Они также определяют тип и цвет ваших волос.Поврежденные волосяные фолликулы могут вызвать воспаление или выпадение волос. Таким образом, понимание науки, лежащей в основе ваших волосяных фолликулов, и выбор правильного режима ухода за волосами имеет решающее значение, если вы хотите здоровые волосы.
Начните с познания своей кожи
Структура и цикл роста волосяных фолликулов
Волосяной фолликул — это структура, похожая на чулок, которая содержит клетки и соединительную ткань и окружает корень волоса. Он существует в дерме и эпидермисе, двух верхних слоях кожи.Для наглядности представьте, что волосяной фолликул — это ваза, а волосы — как стебель цветка.
Изображения героев / Getty Images
Структура волосяных фолликулов проста и понятна, но их функции и цикл роста довольно сложны. Любое существенное изменение нормального цикла роста волосяного фолликула может привести к такому состоянию волос, как очаговая алопеция или телогеновая алопеция.
Структура волосяного фолликула
Волосяные фолликулы состоят из множества различных компонентов, но это четыре ключевые структуры.
Папилла
Сосочек состоит из соединительной ткани и кровеносных сосудов, питающих растущие волосы. Он существует в самом основании волосяного фолликула.
Зародышевый матрикс
В зародышевом матриксе, который также называют «матрицей», клетки производят новые волосы по мере их отмирания и выпадения. Он также расположен в нижней части волосяного фолликула.
Лампа
Луковица представляет собой округлую структуру в форме луковицы в нижней части «чулка» волосяного фолликула, которая окружает сосочек и зародышевый матрикс и питается кровеносными сосудами.Это живая часть волос. На самом деле волосы, которые видны над поверхностью кожи, на самом деле мертвые.
Луковица содержит несколько типов стволовых клеток, которые делятся каждые 23–72 часа, быстрее, чем любые другие клетки в организме. Луковица также содержит гормоны, которые влияют на рост и структуру волос на разных этапах жизни, например, в период полового созревания и во время беременности.
Луковица также содержит гормоны, которые влияют на рост и структуру волос на разных этапах жизни, например, в период полового созревания и во время беременности.
Выпуклость
Область выпуклости расположена в средней части (также известной как перешеек) волосяного фолликула.Он содержит стволовые клетки, которые делятся и регенерируют не только новые волосяные фолликулы, но и сальные железы, и эпидермис.
Выпуклость также служит точкой прикрепления арректирующих пилей — крошечной полоски мышечной ткани. Из-за сокращения этих мышц волосы встают дыбом, когда у вас мурашки по коже.
Цикл роста волос
Скорость роста волос варьируется от человека к человеку, но средняя скорость роста составляет примерно полдюйма в месяц или шесть дюймов в год.Цикл роста волос делится на три фазы: анаген, катаген и телоген. Вот еще несколько подробностей о каждой фазе.
Анаген
Это фаза роста. Фаза анагена в нормальных волосяных фолликулах кожи головы длится примерно от двух до шести лет, тогда как волосы на бровях длится примерно 70 дней в фазе анагена.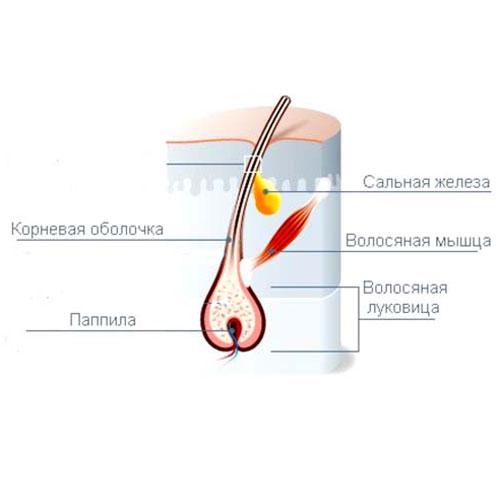
Волосы анагена также различаются по размеру: от длинных толстых терминальных волос до коротких пушковых волос светлого цвета. Повышенный уровень гормонов в период полового созревания превращает пушковые волосы (почти бесцветные) в терминальные (более темные и крупные).Взаимодействие с другими людьми
Катаген
Это фаза регресса. В течение нескольких недель скорость роста волос замедляется, а волосяной фолликул сокращается. Фаза катагена длится две-три недели.
Телоген
Это фаза покоя, которая длится примерно три месяца. Через несколько месяцев волосы перестают расти и отделяются от волосяного фолликула. Новые волосы начинают расти и выталкивают старые мертвые волосы.
В периоды стресса большее количество волос попадает в фазу телогена и начинает выпадать.Люди теряют от 50 до 100 волос на коже головы в день, но стресс может вызвать заметное выпадение волос.
Функциональная регенерация волосяного фолликула: обновленный обзор
Паус, Р. и Фойцик, К. В поисках «часов цикла волос»: экскурсия. Дифференциация 72 , 489–511 (2004).
CAS
PubMed
Статья
Google Scholar
Rezza, A. et al. Сигнальные сети между предшественниками стволовых клеток, предшественниками, усиливающими транзит, и их ниша в развивающихся волосяных фолликулах. Cell Rep. 14 , 3001–3018 (2016).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Ren, X. et al. Делеция Lgr4 задерживает цикл роста волос и подавляет активацию стволовых клеток волосяных фолликулов. J. Invest. Дерматол. 140 , 1706–1712 (2020).
CAS
PubMed
Статья
Google Scholar
Houschyar, K. S. et al. Молекулярные механизмы роста и регенерации волос: современное понимание и новые парадигмы. Дерматология 236 , 271–280 (2020).
PubMed
Статья
Google Scholar
Талавера-Адаме, Д., Ньюман, Д. и Ньюман, Н. Традиционные и новые методы лечения андрогенной алопеции на основе стволовых клеток. Клонирование стволовых клеток 10 , 11–19 (2017).
PubMed
PubMed Central
Google Scholar
Джахода, К. А., Уайтхаус, Дж., Рейнольдс, А. Дж. И Хоул, Н. Дермальные клетки волосяного фолликула дифференцируются на адипогенные и остеогенные клоны. Exp. Дерматол. 12 , 849–859 (2003).
PubMed
Статья
Google Scholar
Driskell, R.R. et al. Четкие клоны фибробластов определяют архитектуру дермы в развитии и восстановлении кожи. Природа 504 , 277–281 (2013).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Миллар С. Э. Молекулярные механизмы, регулирующие развитие волосяных фолликулов. J. Invest. Дерматол. 118 , 216–225 (2002).
CAS
PubMed
Статья
Google Scholar
Хорн К. А., Джахода С. А. и Оливер Р. Ф. Рост усов, индуцированный имплантацией культивированных клеток дермального сосочка вибриссы взрослой крысе. J. Embryol. Exp. Морфол. 97 , 111–124 (1986).
CAS
PubMed
Google Scholar
Jahoda, C.A. et al. Регенерация волосяного фолликула человека после ампутации и пересадки голой мыши. J. Invest. Дерматол. 107 , 804–807 (1996).
CAS
PubMed
Статья
Google Scholar
Кишимото, Дж., Burgeson, R.E. & Morgan, B.A. Передача сигналов Wnt поддерживает индуцирующую волосы активность дермального сосочка. Genes Dev. 14 , 1181–1185 (2000).
CAS
PubMed
PubMed Central
Google Scholar
Джахода, К. А., Хорн, К. А. и Оливер, Р. Ф. Индукция роста волос путем имплантации культивированных клеток дермального сосочка. Nature 311 , 560–562 (1984).
CAS
PubMed
Статья
Google Scholar
Феррарис, К., Бернар, Б. А. и Дуайли, Д. Взрослые эпидермальные кератиноциты наделены способностью формировать волосяной покров. Внутр. J. Dev. Биол. 41 , 491–498 (1997).
CAS
PubMed
Google Scholar
Blanpain, C. et al. Самообновление, мультипотентность и наличие двух популяций клеток в нише эпителиальных стволовых клеток. Cell 118 , 635–648 (2004).
CAS
Статья
PubMed
Google Scholar
Xing, L. & Kobayashi, K. Способность трансплантированного культивированного эпителия реагировать на дермальные сосочки. Tissue Eng. 7 , 535–544 (2001).
CAS
PubMed
Статья
Google Scholar
Ehama, R. et al. Регенерация волосяных фолликулов с использованием привитых клеток грызунов и человека. J. Invest. Дерматол. 127 , 2106–2115 (2007).
CAS
PubMed
Статья
Google Scholar
Zhang, L. et al. Индукция регенерации волосяного фолликула культивированными клетками дермального сосочка мыши в de novo регенерированных тканях кожи. J. Tissue Eng. Regen. Med. 13 , 1641–1650 (2019).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Феррарис, К., Чалоин-Дюфау, С. и Дхуайи, Д. Трансдифференцировка эмбриональных и постнатальных эпителиальных клеток роговицы кролика. Дифференциация 57 , 89–96 (1994).
CAS
PubMed
Статья
Google Scholar
Fliniaux, I., Viallet, J. P., Dhouailly, D. & Jahoda, C. A. B. Трансформация эпителия амниона в кожу и регенерация волосяных фолликулов. Дифференциация 72 , 558–565 (2004).
CAS
PubMed
Статья
Google Scholar
Zhang, M. et al. Предварительные исследования регенерации волосяного фолликула путем инъекции эпидермальных стволовых клеток и клеток дермального сосочка голым мышам. Банк клеточных тканей 21 , 321–327 (2020).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Чжоу, Л.и другие. CD133-положительные лиганды Wnt, происходящие из дермальных сосочков, регулируют послеродовой рост волос. Biochem. J. 473 , 3291–3305 (2016).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Feng, M., Yang, G. & Wu, J. Нацеливание Versican посредством РНК-интерференции подавляет совокупный рост клеток дермального сосочка. Clin. Exp. Дерматол. 36 , 77–84 (2011).
CAS
PubMed
Статья
Google Scholar
Мацузаки Т. и Йошизато К. Роль клеток волосяного сосочка в процессах индукции и регенерации волосяных фолликулов. Регенерация для восстановления ран. 6 , 524–530 (1998).
CAS
PubMed
Статья
Google Scholar
Lu, Z. F. et al. Экспрессия bFGF, ET-1 и SCF в клетках дермального сосочка и связь с их биологическими свойствами. Чжэцзян Да Сюэ Сюэ Бао И Сюэ Бань. 33 , 296–299 (2004).
CAS
PubMed
Google Scholar
Trueb, R.M. Дальнейшие клинические доказательства влияния IGF-1 на рост волос и алопецию. Лыжи. Придатки Disord. 4 , 90–95 (2018).
Артикул
Google Scholar
Na, J. I. et al. Экспрессия гистидиндекарбоксилазы влияет на неофолликулогенез кожных клеток новорожденных мышей. J. Dermatol. Sci. 67 , 95–100 (2012).
CAS
PubMed
Статья
Google Scholar
Liu, S. & Leask, A. CCN2 модулирует цикличность волосяных фолликулов у мышей. Мол. Биол. Ячейка 24 , 3939–3944 (2013).
PubMed
PubMed Central
Статья
Google Scholar
Lim, C.H. et al. Ежик стимулирует регенерацию волосяных фолликулов, создавая индуктивную дерму во время заживления ран на коже мыши. Нат. Commun. 9 , 4903 (2018).
PubMed
PubMed Central
Статья
CAS
Google Scholar
Yu, Z. et al. Hoxc-зависимая гетерогенность мезенхимальных ниш управляет регенерацией региональных волосяных фолликулов. Стволовые клетки клеток 23 , 487–500.e486 (2018).
CAS
PubMed
Статья
Google Scholar
Ли, Ю. Р. и др. Монотерпеноид лолиолид регулирует индукцию волосяного фолликула клеток дермального сосочка человека, активируя сигнальный путь Akt / бета-катенин. J. Microbiol. Biotechnol. 29 , 1830–1840 (2019).
CAS
PubMed
Статья
Google Scholar
Reynolds, A.J. et al. Трансгендерная индукция волосяных фолликулов. Nature 402 , 33–34 (1999).
CAS
PubMed
Статья
Google Scholar
Канг Б. М. и др. Образование сфер увеличивает способность культивируемых клеток кожных сосочков человека индуцировать волосяные фолликулы из эпидермальных клеток мыши в анализе восстановления. J. Invest. Дерматол. 132 , 237–239 (2012).
CAS
PubMed
Статья
Google Scholar
Choi, M. et al. Метаболизм глюкозы регулирует экспрессию индуцирующих волосы генов сфер дермальных сосочков посредством ацетилирования гистонов. Sci. Отчетность 10 , 4887 (2020).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Lin, B.J. et al. LncRNA-XIST способствует индуцированной дермальным сосочком регенерации волосяного фолликула путем нацеливания на miR-424 для активации передачи сигналов hedgehog. Cell Signal. 109623 (2020).
Ahmed, N. S. et al. Эпидермальный E-кадгерин-зависимый путь бета-катенина является фитохимически индуцируемым и ускоряет цикл волос в анагене. Мол. Ther. 25 , 2502–2512 (2017).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Aoi, N. et al. 1Alpha, 25-дигидроксивитамин D3 модулирует способность клеток дермального сосочка индуцировать волосы: терапевтический потенциал для регенерации волос. Stem Cells Пер. Med. 1 , 615–626 (2012).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Miao, Y. et al. Влияние PRP на пролиферацию клеток дермального сосочка и регенерацию волосяного фолликула у мышей. Чжунхуа Чжэн Син Вай Кэ За Чжи. 29 , 131–135 (2013).
PubMed
Google Scholar
Шен, Х., Ченг, Х., Чен, Х. и Чжан, Дж. Идентификация ключевых генов, индуцированных богатой тромбоцитами плазмой в клетках кожных сосочков человека с использованием методов биоинформатики. Мол. Med. Rep. 15 , 81–88 (2017).
CAS
PubMed
Статья
Google Scholar
Su, Y. S. et al. Икариин способствует росту волосяных фолликулов мыши за счет увеличения экспрессии инсулиноподобного фактора роста 1 в папиллярных клетках дермы. Clin. Exp. Дерматол. 42 , 287–294 (2017).
PubMed
Статья
Google Scholar
Инамацу, М., Matsuzaki, T., Iwanari, H. & Yoshizato, K. Создание линий клеток дермального сосочка крысы, которые поддерживают способность индуцировать волосяные фолликулы из афолликулярной кожи. J. Invest. Дерматол. 111 , 767–775 (1998).
CAS
PubMed
Статья
Google Scholar
Райнвальд, Дж. Г. и Грин, Х. Серийное культивирование штаммов эпидермальных кератиноцитов человека: образование ороговевших колоний из отдельных клеток. Cell 6 , 331–343 (1975).
CAS
Статья
PubMed
Google Scholar
Rendl, M., Polak, L. & Fuchs, E. Передача сигналов BMP в клетках дермального сосочка необходима для их индуктивных свойств волосяного фолликула. Genes Dev. 22 , 543–557 (2008).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Osada, A. et al. Долгосрочная культура клеток вибриссального сосочка кожи мыши и индукция волосяного фолликула de novo. Tissue Eng. 13 , 975–982 (2007).
CAS
PubMed
Статья
Google Scholar
Harel, S. et al. Фармакологическое подавление передачи сигналов JAK-STAT способствует росту волос. Sci. Adv. 1 , e1500973 (2015).
PubMed
PubMed Central
Статья
Google Scholar
Toma, J. G. et al. Выделение мультипотентных взрослых стволовых клеток из дермы кожи млекопитающих. Нат. Cell Biol. 3 , 778–784 (2001).
CAS
PubMed
Статья
Google Scholar
Biernaskie, J. et al. SKP происходят из предшественников волосяных фолликулов и проявляют свойства взрослых дермальных стволовых клеток. Стволовые клетки клетки 5 , 610–623 (2009).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Fernandes, K. J. et al. Дермальная ниша для мультипотентных клеток-предшественников, происходящих из кожи взрослых. Нат. Cell Biol. 6 , 1082–1093 (2004).
CAS
PubMed
Статья
Google Scholar
Rahmani, W. et al. Дермальные стволовые клетки волосяного фолликула регенерируют кожную оболочку, повторно заселяют дермальный сосочек и изменяют тип волос. Dev. Ячейка 31 , 543–558 (2014).
CAS
PubMed
Статья
Google Scholar
Guo, L. et al. TSA восстанавливает индуктивную способность волосяных фолликулов предшественников, полученных из кожи. Sci. Отчетность 9 , 2867 (2019).
PubMed
PubMed Central
Статья
CAS
Google Scholar
Gonzalez, R. et al. Передача сигналов тромбоцитарного фактора роста модулирует поддержание и самообновление дермальных стволовых клеток волосяного фолликула. NPJ Regen. Med. 2 , 11 (2017).
PubMed
PubMed Central
Статья
CAS
Google Scholar
Hagner, A. & Biernaskie, J. Выделение и дифференциация дермальных предшественников волосяных фолликулов. Methods Mol. Биол. 989 , 247–263 (2013).
CAS
PubMed
Статья
Google Scholar
Ван, X., Донг, С. и Ву, Y. Выделение и культивирование прекурсоров, полученных из кожи. Methods Mol. Биол. 1879 , 149–152 (2019).
CAS
PubMed
Статья
Google Scholar
Агабалян Н.А. и др. Повышенное размножение и поддержание индуктивной функции клеток-предшественников кожи в биореакторах с перемешиваемой суспензией с компьютерным управлением. Stem Cells Пер. Med. 6 , 434–443 (2017).
CAS
PubMed
Статья
Google Scholar
Охьяма М. Выпуклость волосяного фолликула: удивительный резервуар эпителиальных стволовых клеток. J. Dermatol. Sci. 46 , 81–89 (2007).
CAS
PubMed
Статья
Google Scholar
Hsu, Y. C., Li, L. & Fuchs, E. Клетки, усиливающие транзит, организуют активность стволовых клеток и регенерацию тканей. Ячейка 157 , 935–949 (2014).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Greco, V. et al. Двухэтапный механизм активации стволовых клеток во время регенерации волос. Стволовые клетки клеток 4 , 155–169 (2009).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Hsu, Y. C., Pasolli, H. A. & Fuchs, E. Динамика между стволовыми клетками, нишей и потомством в волосяном фолликуле. Cell 144 , 92–105 (2011).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Rhee, H., Polak, L. & Fuchs, E. Lhx2 поддерживает характер стволовых клеток в волосяных фолликулах. Наука 312 , 1946–1949 (2006).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Vidal, V. P. et al. Sox9 важен для дифференцировки наружной корневой оболочки и формирования компартмента стволовых клеток волос. Curr. Биол. 15 , 1340–1351 (2005).
CAS
PubMed
Статья
Google Scholar
Zhang, S. et al. Стволовые клетки волосяных фолликулов, полученные из одной вибриссы крысы с помощью органной культуры, восстанавливают волосяные фолликулы in vivo. Трансплантация клеток. 21 , 1075–1085 (2012).
PubMed
Статья
Google Scholar
Ge, Y. et al. Микросреда стареющей кожи определяет поведение стволовых клеток. Proc. Natl Acad. Sci. США 117 , 5339–5350 (2020).
CAS
PubMed
Статья
Google Scholar
Beaudoin, G.M., Sisk, J.M., Coulombe, P.A. & Thompson, C.C. Безволосый запускает реактивацию роста волос, стимулируя передачу сигналов Wnt. Proc. Natl Acad. Sci. США 102 , 14653–14658 (2005).
CAS
PubMed
Статья
Google Scholar
Ito, M. et al. Wnt-зависимая регенерация волосяных фолликулов de novo в коже взрослых мышей после ранения. Nature 447 , 316–320 (2007).
CAS
PubMed
Статья
Google Scholar
Li, S. et al. Человеческие TSC2-нулевые фибробластоподобные клетки индуцируют регенерацию волосяного фолликула и морфогенез гамартомы. Нат. Commun. 2 , 235 (2011).
PubMed
PubMed Central
Статья
CAS
Google Scholar
Ohyama, M. et al. Восстановление внутренних свойств кожных сосочков человека in vitro. J. Cell Sci. 125 , 4114–4125 (2012).
CAS
PubMed
Статья
Google Scholar
Veraitch, O. et al. Вызванные человеком эктодермальные клетки-предшественники плюрипотентных стволовых клеток вносят вклад в морфогенез волосяных фолликулов in vivo. J. Invest. Дерматол. 133 , 1479–1488 (2013).
CAS
PubMed
Статья
Google Scholar
Мучкаева И.А. и др. Получение iPS-клеток из клеток дермального сосочка волосяного фоллита человека. Acta Nat. 6 , 45–53 (2014).
CAS
Статья
Google Scholar
Ито, М., Киуру, М., Каир, М. С. и Кристиано, А. М. Генерация кератиноцитов из нормальных и рецессивных дистрофических буллезных эпидермолизов индуцированных плюрипотентных стволовых клеток. Proc. Natl Acad. Sci. США 108 , 8797–8802 (2011).
CAS
PubMed
Статья
Google Scholar
Lim, S.J. et al. Индуцированные плюрипотентные стволовые клетки из кератиноцитов волосяных фолликулов человека как потенциальный источник для клонирования волосяных фолликулов in vitro. PeerJ 4 , e2695 (2016).
PubMed
PubMed Central
Статья
CAS
Google Scholar
Zhao, Q. et al. Химически индуцированная трансформация дермальных фибробластов человека в дермальные сосочковые клетки, вызывающие рост волос. Cell Prolif. 52 , e12652 (2019).
PubMed
PubMed Central
Статья
Google Scholar
Wenzel, V. et al. Наивные взрослые стволовые клетки пациентов с синдромом прогерии Хатчинсона-Гилфорда экспрессируют низкие уровни прогерина in vivo. Biol. Открыть 1 , 516–526 (2012).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Budel, L. & Djabali, K. Быстрая изоляция и размножение кожных клеток-предшественников из первичных культур фибробластов человека. Biol. Открыть 6 , 1745–1755 (2017).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Liu, S. et al. Путь PI3K-Akt ингибирует старение и способствует самообновлению предшественников, происходящих из кожи человека, in vitro. Ячейка старения 10 , 661–674 (2011).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Hou, P. et al. Плюрипотентные стволовые клетки, индуцированные из соматических клеток мыши низкомолекулярными соединениями. Наука 341 , 651–654 (2013).
CAS
PubMed
Статья
Google Scholar
Lim, X. et al. Axin2 маркирует покоящиеся стволовые клетки выпуклости волосяного фолликула, которые поддерживаются аутокринной передачей сигналов Wnt / beta-catenin. Proc. Natl Acad. Sci. США 113 , E1498–1505 (2016).
CAS
PubMed
Статья
Google Scholar
Ito, M., Kizawa, K., Hamada, K. & Cotsarelis, G. Стволовые клетки волосяного фолликула в нижней выпуклости образуют вторичный зародыш, биохимически отличную, но функционально эквивалентную популяцию клеток-предшественников, в нижнем выступе. прекращение катагена. Дифференциация 72 , 548–557 (2004).
Артикул
PubMed
Google Scholar
Taylor, G. et al. Участие фолликулярных стволовых клеток в формировании не только фолликула, но и эпидермиса. Cell 102 , 451–461 (2000).
CAS
PubMed
Статья
Google Scholar
Paus, R. et al. Нейроэндокринология волосяного фолликула: принципы и клинические перспективы. Trends Mol. Med. 20 , 559–570 (2014).
CAS
PubMed
Статья
Google Scholar
Schneider, M. R., Schmidt-Ullrich, R. & Paus, R. Волосяной фолликул как динамический миниорган. Curr. Биол. 19 , R132–142 (2009).
CAS
PubMed
Статья
Google Scholar
Chen, Y. et al. Сигнальный путь PI3K / Akt важен для регенерации волосяных фолликулов de novo. Stem Cell Res. Ther. 11 , 144 (2020).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Matsumura, H. et al. Старение волосяного фолликула происходит за счет трансэпидермального удаления стволовых клеток посредством протеолиза COL17A1. Наука 351 , aad4395 (2016).
Артикул
CAS
PubMed
Google Scholar
Nakao, K. et al. Разработка метода биоинженерных зародышей органов. Нат. Методы 4 , 227–230 (2007).
CAS
PubMed
Статья
Google Scholar
Asakawa, K. et al. Регенерация органов волос посредством трансплантации биоинженерных фолликулярных единиц. Sci. Отчет 2 , 424 (2012).
PubMed
PubMed Central
Статья
CAS
Google Scholar
Toyoshima, K. E. et al. Полнофункциональная регенерация волосяных фолликулов за счет перестройки стволовых клеток и их ниш. Нат. Commun. 3 , 784 (2012).
PubMed
PubMed Central
Статья
CAS
Google Scholar
Fan, S. M. et al. Стимулирование регенерации волосяных фолликулов с помощью секретируемых белков, обогащенных эмбриональной кожей. Биоматериалы 167 , 121–131 (2018).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Морган, Б. А. Дермальный сосочек: поучительная ниша для эпителиальных стволовых и предшественников клеток в развитии и регенерации волосяного фолликула. Cold Spring Harb. Перспектива. Med. 4 , а015180 (2014).
PubMed
PubMed Central
Статья
CAS
Google Scholar
Коллинз, К. А., Кречмар, К. и Ватт, Ф. М. Перепрограммирование дермы взрослого человека в неонатальное состояние посредством эпидермальной активации бета-катенина. Разработка 138 , 5189–5199 (2011).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Yang, R. et al. Прямое преобразование фибробластов мыши и человека в функциональные меланоциты определенными факторами. Нат. Commun. 5 , 5807 (2014).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Kim, H. S. et al. Современные системы доставки лекарств и искусственные кожные трансплантаты для заживления кожных ран. Adv. Препарат Делив. Ред. 146 , 209–239 (2019).
CAS
PubMed
Статья
Google Scholar
Zhou, L. et al. Активация передачи сигналов бета-катенином в CD133-положительных клетках сосочка дермы увеличивает индуктивность волос. FEBS J. 283 , 2823–2835 (2016).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Dong, L. et al. Wnt1a поддерживает характеристики клеток дермальных сосочков, которые индуцируют регенерацию шерсти мышей в системе трехмерного прекультивирования. J. Tissue Eng. Regen. Med. 11 , 1479–1489 (2017).
CAS
PubMed
Статья
Google Scholar
Young, T.H. et al. Самосборка клеток дермального сосочка в индуктивные сфероидальные микроткани на мембранах из поли (этилен-винилового спирта) для регенерации волосяного фолликула. Биоматериалы 29 , 3521–3530 (2008).
CAS
PubMed
Статья
Google Scholar
Huang, Y.C. et al. Масштабируемое производство контролируемых сфероидов дермальных сосочков на поверхности PVA и влияние размера сфероидов на регенерацию волосяных фолликулов. Биоматериалы 34 , 442–451 (2013).
CAS
PubMed
Статья
Google Scholar
Lin, B. et al. Висячая капля, управляемая поверхностным натяжением: создание управляемого трехмерного сфероида из высокопроходных клеток дермального сосочка человека и формирование индуктивных микротканей для регенерации волосяных фолликулов. ACS Appl. Матер. Интерфейсы 8 , 5906–5916 (2016).
CAS
PubMed
Статья
Google Scholar
Kwack, M.H. et al. Экзосомы, полученные из клеток кожных сосочков человека, способствуют росту волос в культивируемых волосяных фолликулах человека и увеличивают способность культивированных сфер кожных сосочков к волосам. Exp. Дерматол. 28 , 854–857 (2019).
CAS
PubMed
Статья
Google Scholar
Hu, S. et al. Кожные экзосомы, содержащие miR-218-5p, способствуют регенерации волос, регулируя передачу сигналов бета-катенина. Sci. Adv. 6 , eaba1685 (2020).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Wang, J. et al. Наноинкапсуляция снизу вверх от отдельных клеток до настраиваемых и масштабируемых клеточных сфероидов для регенерации волосяных фолликулов. Adv. Здоровьеc. Mater 7 , 1–9 (2018).
Артикул
CAS
Google Scholar
Zhang, X. et al. Использование гидрогеля внеклеточного матрикса из плаценты человека для восстановления волосяного потенциала клеток дермального сосочка. Regen. Med. 14 , 741–751 (2019).
CAS
PubMed
Статья
Google Scholar
Vahav, I. et al. Реконструированная человеческая кожа показывает эпидермальную инвагинацию в сторону интегрированных неопапилл, что указывает на раннее формирование волосяных фолликулов in vitro. J. Tissue Eng.Regen. Med. 14 , 761–773 (2020).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Zhang, K. et al. Клеточная структура нановолокон с характеристиками, способствующими секреторной активности, для образования многоклеточных сфероидов и регенерации волосяных фолликулов. ACS Appl. Матер. Интерфейсы 12 , 7931–7941 (2020).
CAS
PubMed
Статья
Google Scholar
Abaci, H.E. et al. Тканевая инженерия человеческих волосяных фолликулов с использованием биомиметического подхода к развитию. Нат. Commun. 9 , 5301 (2018).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Wang, X. et al. Самособирающиеся каркасы из пептидного гидрогеля поддерживают регенерацию волосяных фолликулов на основе стволовых клеток. Наномедицина 12 , 2115–2125 (2016).
CAS
PubMed
Статья
Google Scholar
Wang, X. et al. Регенерация волосяного фолликула и сальной железы de novo культивированными эпидермальными стволовыми клетками и предшественниками кожного происхождения. Stem Cells Пер. Med. 5 , 1695–1706 (2016).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Ellis-Behnke, R.G. et al. Нано нейровязание: каркас из пептидных нановолокон для восстановления мозга и регенерации аксонов с функциональным возвращением зрения. Proc. Natl Acad. Sci. США 103 , 5054–5059 (2006).
CAS
PubMed
Статья
Google Scholar
Meng, H. et al. Эффект самособирающегося каркаса из пептидных нановолокон (пептид) при использовании в качестве перевязочного материала для лечения глубоких ожогов второй степени у крыс. J. Biomed. Матер. Res. B 89 , 379–391 (2009).
Артикул
CAS
Google Scholar
Kumada, Y. & Zhang, S. Значительное производство коллагена типа I и типа III из фибробластов периодонтальной связки человека в трехмерных пептидных каркасах без дополнительных факторов роста. PLoS ONE 5 , e10305 (2010).
PubMed
PubMed Central
Статья
CAS
Google Scholar
Carrejo, N.C. et al. Многодоменный пептидный гидрогель ускоряет заживление полнослойных ран у мышей с диабетом. ACS Biomater. Sci. Англ. 4 , 1386–1396 (2018).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Zhang, Y. et al. Многофункциональные полые мезопористые мезопористые мембраны из диоксида кремния / поликапролактона, легированные цинком, с усиленной регенерацией волосяных фолликулов и антибактериальной активностью для заживления ран. Наноразмер 11 , 6315–6333 (2019).
CAS
PubMed
Статья
Google Scholar
Bader, A. et al. Регенерация кожи с конической структурой и структурой волосяного фолликула при глубоких ожоговых повреждениях второй степени за счет комбинированной экспрессии рецептора EPO и общего рецептора бета путем локальной подкожной инъекции наноразмерного rhEPO. Внутр. J. Nanomed. 7 , 1227–1237 (2012).
CAS
Google Scholar
Giri, P. et al. Регенерация кожи при глубоких ожогах второй степени с помощью инфузионного насоса или местного применения геля рекомбинантного человеческого эритропоэтина. Drug Des. Dev. Ther. 9 , 2565–2579 (2015).
CAS
Статья
Google Scholar
Zhang, Z. et al. Дизайн многофункционального биоматериала, вдохновленный древней китайской медициной, для восстановления волос на обожженной коже. ACS Appl. Матер. Интерфейсы 12 , 12489–12499 (2020).
PubMed
Статья
CAS
Google Scholar
Кливерс, Х. Моделирование развития и болезней с помощью органоидов. Ячейка 165 , 1586–1597 (2016).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Gupta, A.C. et al. Создание in vitro органоидной модели дермального сосочка волосяного фолликула человека. J. Cell. Physiol. 233 , 9015–9030 (2018).
CAS
PubMed
Статья
Google Scholar
Lee, J. et al. Развитие волосяных фолликулов в органоидах кожи, полученных из плюрипотентных стволовых клеток мыши. Cell Rep. 22 , 242–254 (2018).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Su, Y. et al. Предварительная агрегация дермальных и эпидермальных стволовых клеток-предшественников кожи головы активирует путь WNT и способствует образованию волосяных фолликулов в системах in vitro и in vivo. Stem Cell Res.Ther. 10 , 403 (2019).
PubMed
PubMed Central
Статья
Google Scholar
Takagi, R. et al. Биоинженерия трехмерной системы покровных органов из iPS-клеток с использованием модели трансплантации in vivo. Sci. Adv. 2 , e1500887 (2016).
PubMed
PubMed Central
Статья
CAS
Google Scholar
Стенн, К. С. и Паус, Р. Контроль цикличности волосяных фолликулов. Physiol. Ред. 81 , 449–494 (2001).
CAS
PubMed
Статья
Google Scholar
Хеберт, Дж. М., Розенквист, Т., Гоц, Дж. И Мартин, Г. Р. FGF5 как регулятор цикла роста волос: данные целевых и спонтанных мутаций. Cell 78 , 1017–1025 (1994).
CAS
PubMed
Статья
Google Scholar
Палмер, Х. Г., Мартинес, Д., Кармелиет, Г. и Ватт, Ф. М. Рецептор витамина D необходим для развития цикла волос у мышей, но не для поддержания компартмента эпидермальных стволовых клеток. J. Invest. Дерматол. 128 , 2113–2117 (2008).
CAS
PubMed
Статья
Google Scholar
Watt, F. M., Estrach, S. & Ambler, C. A. Передача сигналов эпидермальных Notch: дифференциация, рак и адгезия. Curr. Opin. Cell Biol. 20 , 171–179 (2008).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Suen, W. J., Li, S. T. и Yang, L. T. Hes1 регулирует инициирование анагена и регенерацию волосяного фолликула посредством модуляции передачи сигналов hedgehog. Стволовые клетки 38 , 301–314 (2020).
CAS
PubMed
Google Scholar
Legrand, J. M. D. et al. Активация STAT5 в дермальном сосочке важна для индукции фазы роста волосяного фолликула. J. Invest. Дерматол. 136 , 1781–1791 (2016).
CAS
PubMed
Статья
Google Scholar
Watabe, R. et al. Лептин контролирует цикличность волосяных фолликулов. Exp. Дерматол. 23 , 228–229 (2014).
CAS
PubMed
Статья
Google Scholar
Сумикава Ю., Инуи С., Накадзима Т. и Итами С. Контроль цикла волос с помощью лептина как нового индуктора анагена. Exp. Дерматол. 23 , 27–32 (2014).
CAS
PubMed
Статья
Google Scholar
Nakajima, T. et al. Роль MED1 в покое стволовых клеток волосяного фолликула и поддержании нормального цикла волос. J. Invest. Дерматол. 133 , 354–360 (2013).
CAS
PubMed
Статья
Google Scholar
Zhu, K., Xu, C., Liu, M. и Zhang, J. Hairless контролирует решение судьбы волос посредством передачи сигналов Wnt / beta-catenin. Biochem. Биофиз. Res. Commun. 491 , 567–570 (2017).
CAS
PubMed
Статья
Google Scholar
Saini, V. et al. Отсутствие подавления PPARgamma, опосредованного рецептором витамина D (VDR), вызывает алопецию у мышей, не имеющих VDR. FASEB J. 31 , 1059–1066 (2017).
CAS
PubMed
Статья
Google Scholar
Mercati, F. et al. Эпителиальная экспрессия гормона лептина в коже крупного рогатого скота. Eur. J. Histochem. 63 , 9–16 (2019).
Артикул
CAS
Google Scholar
Тонг, X. & Coulombe, P.A. Кератин 17 модулирует циклическое движение волосяных фолликулов TNF-альфа-зависимым образом. Genes Dev. 20 , 1353–1364 (2006).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Zhou, L. et al. Декорин способствует пролиферации и миграции кератиноцитов ПРС и поддерживает анаген волос у мышей. Exp. Дерматол. 27 , 1237–1244 (2018).
CAS
PubMed
Статья
Google Scholar
Qiu, W. et al. Hoxc13 является важным регулятором цикла мышей. Cell Tissue Res. 364 , 149–158 (2016).
CAS
PubMed
Статья
Google Scholar
Jing, J. et al. Экспрессия декорина на протяжении всего цикла волосяного фолликула мыши: зависимость цикла волос и продление фазы анагена. Exp. Дерматол. 23 , 486–491 (2014).
CAS
PubMed
Статья
Google Scholar
Wu, X. J. et al. Экспрессия и расположение фосфо-Artemis (Serine516) в волосяных фолликулах во время индуцированного роста волос мыши. Arch. Дерматол. Res. 304 , 319–324 (2012).
CAS
PubMed
Статья
Google Scholar
Самуэлов Л. и др. Р-кадгерин регулирует рост и цикл человеческого волоса посредством канонической передачи сигналов Wnt и трансформирующего фактора роста-бета2. J. Invest. Дерматол. 132 , 2332–2341 (2012).
CAS
PubMed
Статья
Google Scholar
Семенова Е. и др. Избыточная экспрессия mIGF-1 в кератиноцитах улучшает заживление ран и ускоряет образование волосяных фолликулов и цикличность у мышей. Am. J. Pathol. 173 , 1295–1310 (2008).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Бикле, Д.D. et al. Развитие и прогрессирование алопеции у мышей, у которых отсутствует рецептор витамина D. J. Cell. Physiol. 207 , 340–353 (2006).
CAS
PubMed
Статья
Google Scholar
Пантелеев А.А. и др. К определению патогенеза безволосого фенотипа. J. Invest. Дерматол. 110 , 902–907 (1998).
CAS
PubMed
Статья
Google Scholar
Potter, G. B. et al. Ген безволосости, мутировавший при врожденном облысении, кодирует новый корепрессор ядерного рецептора. Genes Dev. 15 , 2687–2701 (2001).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Al-Nuaimi, Y. et al. Встреча двух хронобиологических систем: циркадные белки Period1 и BMAL1 модулируют часы цикла человеческого волоса. J. Invest.Дерматол. 134 , 610–619 (2014).
CAS
PubMed
Статья
Google Scholar
Castela, M. et al. Передача сигналов Igf1r влияет на переход от анагена к катагену в цикле волос. Exp. Дерматол. 26 , 785–791 (2017).
CAS
PubMed
Статья
Google Scholar
Bichsel, K. J. et al. Рецептор эпидермального фактора роста снижает Stathmin 1 и запускает катаген у мышей. Exp. Дерматол. 25 , 275–281 (2016).
CAS
PubMed
Статья
Google Scholar
Колсон-Томас, В. Дж., Гестейра, Т. Ф., Эско, Дж. И Као, В. Гепарансульфат регулирует морфогенез и гомеостаз волосяного фолликула и сальных желез. J. Biol. Chem. 289 , 25211–25226 (2014).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Kim, B. K. et al. Повышенная экспрессия Dkk1 с помощью HR связана с изменением цикла волос у мышей с плохим волосяным покровом. J. Dermatol. Sci. 74 , 81–87 (2014).
CAS
PubMed
Статья
Google Scholar
Kwack, M. H., Kim, M. K., Kim, J. C. & Sung, Y. K. Dickkopf 1 способствует регрессии волосяных фолликулов. J. Invest. Дерматол. 132 , 1554–1560 (2012).
CAS
PubMed
Статья
Google Scholar
Kandyba, E. & Kobielak, K. Wnt7b является важным внутренним регулятором гомеостаза стволовых клеток волосяного фолликула и круговорота волосяного фолликула. Стволовые клетки 32 , 886–901 (2014).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Bai, X. et al. Роль GasderminA3 в переходе между катагеном и телогеном во время круговорота волос. J. Invest. Дерматол. 135 , 2162–2172 (2015).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Plikus, M. V. et al. Циклическая передача сигналов дермального BMP регулирует активацию стволовых клеток во время регенерации волос. Nature 451 , 340–344 (2008).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Wu, P. et al. Баланс Bmp6 и Wnt10b регулирует переход телоген-анаген волосяных фолликулов. Cell Commun. Сигнал. 17 , 16 (2019).
PubMed
PubMed Central
Статья
Google Scholar
Calvo-Sanchez, M. I. et al. Роль корецептора Tgf-beta / Bmp Endoglin в молекулярном осцилляторе, который регулирует цикл волосяного фолликула. J. Mol. Клетка. Биол. 11 , 39–52 (2019).
CAS
PubMed
Статья
Google Scholar
Krieger, K. et al. NF-kappaB участвует в контроле цикла волосяного покрова мышей и играет различные роли в различных типах волосяных фолликулов шерсти. J. Invest. Дерматол. 138 , 256–264 (2018).
CAS
PubMed
Статья
Google Scholar
Фолликулит: виды, лечение
Обзор
Что такое фолликулит?
Фолликулит — распространенное заболевание, которое часто встречается на лице, руках, верхней части спины и голени.Существует много различных типов, но все они характеризуются воспалением (отеком), инфекцией или раздражением волосяного фолликула (пространство в коже, в котором находится корень волоса и сальные железы). Когда эти фолликулы инфицируются или раздражаются, могут развиться покраснение, раздражение, зуд и неровности.
Какие бывают типы фолликулита?
К типам фолликулита относятся следующие:
- Staphylococcus aureus фолликулит: Инфекция волосяного фолликула бактериями Staphylococcus aureus является одной из наиболее частых причин фолликулита.На коже можно увидеть небольшие красные или белые прыщики, заполненные гноем. Пораженный участок часто проходит (выздоравливает) в течение нескольких дней, и за ним можно ухаживать дома. Однако в тяжелых случаях фолликулит следует лечить у медицинского работника.
- Pseudomonas aeruginosa («горячая ванна») фолликулит: Pseudomonas aeruginosa — это бактерия, которая размножается в нагретой движущейся воде (горячие ванны, водовороты, водные горки). Он может заразить волосяной фолликул и вызвать сыпь, очень похожую на вызванную стафилококками.Иногда сыпь вызывает зуд. Фолликулит в гидромассажной ванне обычно проходит сам по себе в течение нескольких дней, но некоторым пациентам может потребоваться медицинская помощь.
- Malassezia фолликулит : Malassezia — это семейство дрожжей, которые обычно встречаются на коже. Иногда, когда Malassezia попадает в волосяные фолликулы, это может вызвать зуд, который выглядит как высыпание прыщей, обычно в верхней части груди и спине. Эта форма фолликулита усугубляется потоотделением.Часто помогает ежедневное мытье пораженных участков кожи шампунем от перхоти.
- Барбадосский псевдофолликулит : Также называемый «бритвенными шишками» Барбадосский псевдофолликулит обычно возникает в области бороды. После стрижки волос на бороде бритвой резко остриженные края могут снова превратиться в кожу, вызывая раздражение. Барбадосский псевдофолликулит чаще встречается у людей с вьющимися волосами, особенно у афроамериканских мужчин.
- Барбадосикоз : Барбадосикоз — тяжелая форма фолликулита, связанного с бритьем, с потенциально рубцеванием. Инфицирован весь волосяной фолликул, в результате чего образуются большие красные пустулы.
- Грамотрицательный фолликулит : грамотрицательный фолликулит может развиться после длительного приема антибиотиков для лечения акне. Со временем устойчивые бактерии растут и размножаются, что приводит к обострению прыщей.
- Фурункулы (фурункулы) : Фурункулы или фурункулы возникают при глубоком инфицировании волосяного фолликула.Фурункул часто бывает красным, нежным и болезненным. Через несколько дней болезнь вырастет, и после нее может остаться шрам.
- Карбункулы: Карбункул образуется при появлении нескольких фурункулов в одном месте. Карбункулы обычно больше по размеру и представляют собой комбинацию нескольких инфицированных волосяных фолликулов.
- Эозинофильный фолликулит : Это состояние обычно наблюдается у пациентов с подавленным иммунитетом (иммунная система функционирует не полностью). Есть также форма, которая наблюдается у младенцев.Эозинофильный фолликулит не заразен. Для него характерны зудящие пустулы, чаще всего на плечах, плечах, шее и лбу.
Ведение и лечение
Как лечить и лечить фолликулит?
Некоторые виды фолликулита можно лечить в домашних условиях следующими способами:
- Антибактериальные очищающие средства помогают очистить кожу, ограничивая количество бактерий на коже.
- Теплые полотенца также успокаивают раздраженную кожу.
- Кремы от зуда могут уменьшить дискомфорт.
- Менее тяжелые случаи фолликулита, такие как Pseudomonas folliculitis (сыпь в горячей ванне), обычно проходят в течение нескольких дней без лечения.
В более тяжелых случаях может потребоваться пероральный антибиотик. Более глубокие инфекции, такие как фурункулы и карбункулы, могут потребоваться врачу.
Поскольку Pseudofolliculitis barbae и Sycosis barbae сильно влияют на область бороды, вы можете уменьшить их частоту с помощью:
- Смягчение волос горячей водой перед бритьем.
- Бритье по волосам, а не против них.
- Использование геля или крема для бритья.
- Бритье через день, а не ежедневно.
- Использование электрической бритвы или средства для удаления волос вместо обычного лезвия.
- Избегать натягивания кожи во время бритья.
Профилактика
Как предотвратить другие формы фолликулита?
Инфекции фолликулита в основном связаны с попаданием бактерий, микробов и дрожжей в волосяные фолликулы.Вы можете предотвратить или уменьшить последствия фолликулита с помощью:
- Сохранение кожи в чистоте.
- Уход во время бритья.
- Проверка уровня химических дезинфицирующих средств в гидромассажных ваннах и бассейнах с подогревом перед их использованием.
- Смыть и снять купальник, когда вы выходите из гидромассажной ванны.
- Носить дышащую одежду, чтобы пот не попадал между одеждой и кожей.
Жить с
Когда мне следует позвонить своему врачу по поводу фолликулита?
Хотя во многих случаях фолликулит не является серьезным заболеванием и проходит в течение короткого времени, в тяжелых случаях может потребоваться медицинское лечение.Если фолликулит распространяется, или если есть твердые, болезненные или дренирующие шишки, как можно скорее обратитесь к врачу.
Многомасштабная модель волосяных фолликулов выявляет неоднородные домены, управляющие быстрым пространственно-временным паттерном роста волос
Основные комментарии, которые следует учесть в новой редакции рукописи:
1) В целом, рецензенты считали, что эти результаты следует рассматривать в более широком контексте с собственными работами авторов и другими.Во-первых, в чем разница между этой рукописью, Plikus et al., 2011, и взаимодействием между фолликулами, которое недавно исследовали Chen et al., 2015? Авторы проводят некоторые сравнения в разделе результатов (подраздел «Модель показывает, что кожа является неоднородной возбудимой средой»), но такая логика должна входить в экспериментальный план для введения и результатов. Во-вторых, акцент на передаче сигналов Wnt и Bmp кажется излишним, судя по предыдущим исследованиям нескольких лабораторий. В-третьих, авторы должны обсудить, как подобный подход может помочь в изучении других тканей?
Благодарим рецензентов за предложения.В этом документе мы внесли изменения во Введение и Обсуждение, чтобы уточнить, что эта работа является первой, исследующей регенерацию волос на большей части кожи тела мыши, и ее результаты представляют собой значительный прогресс по сравнению с предыдущими знаниями о поведении роста волос на групповом уровне в спинная кожа. С точки зрения моделирования, это первая многомасштабная модель волосяного фолликула, которая объединяет молекулярную динамику и физический рост в одну модель, состоящую из трех измерений в пространстве, и которая точно предсказывает циклическое поведение на уровне как отдельного волосяного фолликула, так и его состояния. популяции волос в совокупности.С помощью экспериментов и моделирования наша работа показала, что:
a) Физиологические темпы регенерации волос различаются в зависимости от анатомических участков. Это заметно быстрее на подбородке и коже брюшины по сравнению с кожей спины и очень медленно на коже ушей. Таким образом, кожа в целом функционирует как сложный регенерирующий ландшафт с участками быстрого, медленного и очень медленного обновления волос. Наши новые данные в этой редакции (Приложение 1 — рис. 23) показывают, что при таком поведении образуется шуба с переменной густотой волос, которая, вероятно, играет адаптивную роль.В предыдущих работах на коже спины не учитывалось, что динамика регенерации волос заметно различается между участками тела. Теперь мы подчеркнем это во введении и в обсуждении.
b) Сигнальная пара активатор / ингибитор WNT / BMP модулирует регенерацию волос во всех областях кожи. Мы считаем этот вывод важным и не дублирующим предыдущие работы. Наша текущая работа показывает, что передача сигналов WNT / BMP — это универсальный «молекулярный язык» для роста волос на всем теле, а не только на определенном участке тела, таком как кожа спины, показанная в предыдущих работах.Действительно, выбор кожи спины в качестве модели для изучения роста волос в большинстве предшествующих работ в первую очередь обусловлен удобством. Изучая рост волос в других, «менее удобных» участках тела, мы видим, что механизм, управляемый сигнальной парой WNT / BMP, является общим. В этой редакции мы приложили дополнительные усилия для дальнейшего усиления этого важного аспекта нашей работы, предоставив новые данные об активности репортеров WNT и BMP в масштабе всей кожи (рис. 4G и 4I; Приложение 1 — рис. 21), а также новые данные об изменениях в паттерны роста волос у мышей с мутантами WNT и BMP (рис. 4J и 4K; приложение 1 — рис. 22).
c) Несколько дополнительных выводов вытекают непосредственно из вышеизложенных выводов. Во-первых, поскольку все области кожи имеют общий «язык» WNT / BMP, связь между волосами естественным образом возникает через анатомические границы. Альтернативой этому механизму были бы отдельные, сайт-специфические сигналы и полная автономия регионального роста волос. Поскольку связь на основе WNT / BMP возможна через границы, она обеспечивает новое поведение роста волос, не очевидное из предыдущих работ — быстро меняющиеся участки кожи (например, кожа подбородка) действуют как своего рода кардиостимулятор для роста волос по всей коже.Мы подчеркиваем это во Введении и в Обсуждении.
d) Во-вторых, мы обнаружили, что некожные ткани с выраженными профилями экспрессии WNT / BMP могут действовать как сигнальная макросреда для волосяных фолликулов и сильно влиять на скорость их регенерации. В этом исследовании мы показываем, что в ухе мыши комплекс хрящ / мышцы с низким уровнем WNT / высоким BMP замедляет рост волос на коже уха практически до полной остановки. Это открытие расширяет репертуар тканей с функцией сигнальной макросреды, включая близко расположенные анатомические структуры за пределами типичных популяций клеток кожи.Мы выделяем это в Обсуждении.
e) Мы полагаем, что некоторые из недавно обнаруженных свойств регенерации волос могут иметь аналоги в других тканях и органах. Например, доминирующие анатомически определенные кардиостимуляторы распространены в электрически связанных мышечных тканях. В частности, сердечная ткань содержит узлы кардиостимулятора, которые генерируют доминирующие электрические ритмы, управляющие сократительной активностью всего сердца. Подобный кардиостимулятор существует в желудке (зона кардиостимулятора желудка), который контролирует его скоординированную перистальтику.Другие активно регенерирующие органы, такие как желудочно-кишечный тракт и костный мозг, вероятно, содержат анатомические области более быстрой и медленной регенерации, и, возможно, они могут работать вместе, чтобы работать согласованно. Знания, полученные из кожной системы, могут направлять поиск регенеративных ландшафтов в этих и других органах. Поскольку принципы координации, наблюдаемые в коже, могут быть универсальными для крупномасштабных систем формирования паттернов, вероятность того, что они действуют в других органах, высока, несмотря на заметные анатомические различия.Мы выделяем это в Обсуждении.
2) Часто текст написан непонятно. Например, фраза «мы масштабировали модель как в двух (Приложение 2 — рис. 4A), так и в трех измерениях (Приложение 2 — рис. 4B)» не имеет для меня смысла. Отсутствие ясности делает практически невозможным интерпретацию рисунков 1 и 2.
Приносим извинения за недоразумение. В эту редакцию мы включили расширенное описание моделирования как в основной текст (раздел «Результаты»), так и в условные обозначения основных рисунков 1 и 2 и Приложения 2 — рисунок 4.Мы надеемся, что это поможет улучшить ясность моделирования аспектов исследования.
3) Фигура 2A и B не демонстрируют четко «асинхронность от шеи к хвосту» в морфогенезе волос. Имеют ли авторы в виду, что центр отличается от левого края правого?
В самом деле, как отмечают обозреватели, асинхронность от шеи к хвосту в морфогенезе дорсальных волос в точке P0-P1 довольно тонка. Вдобавок наблюдаются тонкие различия между сторонами и центром спинной кожи.Теперь мы явно отметили это в исправленном тексте (раздел «Результаты»). Наши данные на основных рисунках 3 и 4 показывают, что эти начальные пространственно-временные асинхронности в морфогенезе дорсальных волос эффективно «стираются» из-за доминантного роста волос от подбородка / вентрального к дорсальному, распространяющегося в начале второго анагена. Тем не менее, мы указали на эту начальную асинхронность.
На рис. 2F авторы продемонстрировали сценарий того, как может возникнуть билатерально-симметричный паттерн роста волос.Даже с предполагаемыми параметрами, описанными в тексте, неясно, почему распространение анагена из дорсально-вентральной части происходит только в задней части вентральной кожи, например, с t = 50 до t = 62. Предполагается ли, что передне-задние части кожи брюшины тоже разные? Независимо от того, так ли это здесь, авторам следует расширить, чтобы объяснить это явление, потому что оно влияет на результат всей модели. В этой рукописи авторы не обращали внимания на передне-заднюю разницу кожи, но, учитывая характер этого исследования, было бы полезно прояснить, почему авторы решили сделать это.
Мы согласны с критикой рецензентов о том, что результат моделирования на исходном рисунке 2F не полностью соответствует экспериментально наблюдаемым моделям роста волос. При повторном представлении мы критически изучили нашу модель путем всестороннего исследования параметров и обнаружили, что эта функция была результатом определенного набора значений параметров, выбранных для наших исходных симуляций. В этой редакции мы протестировали более широкий диапазон значений параметров и обнаружили, что дорсальное / вентральное распространение не является исключительной особенностью задней части кожи, но может происходить по всей длине дорсальной / вентральной границы.Обновленный набор значений параметров включает: (a) более раннее начало морфогенеза вентрального волосяного фолликула, (b) более низкий порог передачи сигналов для инициации анагена (Δ + = 2,6), (c) более низкий сигнальный шум активатора / ингибитора (λ A = λ I = 0,02), и (d) меньшая длина временного шага (dt = 0,01). Обновленные результаты теперь показаны на основном рисунке 2F, а используемые параметры показаны в Приложении 2 (подраздел «Детали 2D и 3D моделирования». 1 и Приложение 2 — таблицы с 1 по 3. Кроме того, для единообразия, мы повторно — запустите другие модели и обновите Приложение 2 — рисунки 7, 12, 13B, 15–18 и 28–30.Во всех случаях мы получили качественно похожие результаты, которые не меняют наших первоначальных выводов. Наконец, мы также внесли три дополнительных изменения в представление результатов моделирования в файле Приложения 2:
1) Влияние максимальной длины роста волосяного фолликула h max на поведение распространения волн было представлено в дополнительной таблице в исходной документации. Теперь они показаны в Приложении 2 — рисунок 27.
2) Влияние всех доступных рецепторов R A и R I на вентрально-дорсальные волновые взаимодействия было показано в видеороликах в исходной заявке.Теперь они показаны в Приложении 2 — рисунки с 28 по 30.
3) Новое Приложение 2 — таблица 2 добавлена к Приложению 2 — таблица 1. В нем подробно описаны значения параметров, используемые в каждом моделировании.
4) На рисунке 3 используется репортер Wnt для изучения цикличности волос, но неясно, является ли сигнал люциферазы специфичным для кожи.
В исходном исследовании мышей-репортеров Flash WNT, проведенном Hodgson et al., 2014, было тщательно изучено происхождение сигнала люминесценции и сделан вывод, что он специфичен для кожи.В своей работе авторы заявляют, что ‘ […] При вскрытии не было обнаружено сигнала, превышающего уровень WT в органах, кроме кожи как при коротком, так и при длительном времени воздействия, хотя экспрессия люциферазы могла быть обнаружена на уровне РНК, что указывает на то, что активность трансгена на высоком и детектируемом уровне характерна исключительно для кожи без контаминации сигналами внутренних органов […] ‘. Таким образом, мы интерпретируем репортерный сигнал Flash в текущем исследовании как специфичный для кожи. В пересмотренной версии мы указываем на специфическую для кожи природу сигнала Flash и цитируем приведенную выше ссылку.
Результат в Приложении 1 очень интересен. Это говорит о том, что кожа подбородка имеет более быструю цикличность, вместо того, чтобы полагаться на соседние ткани подбородка, такие как хрящ в ухе. Чтобы понять суть, было бы полезно провести Flash-анализ на подбородочном трансплантате и сравнить динамику передачи сигналов Wnt в трансплантате по сравнению с нативной кожей подбородка. Также этот результат следует перенести на основной рисунок, поскольку он служит веским основанием для проведения последующего анализа последовательности РНК.
Мы согласны с тем, что результаты трансплантации поддерживают внутренне более быструю базовую динамику передачи сигналов WNT и / или BMP в коже подбородка. Как и предполагалось, мы рассмотрели эксперименты по пересадке кожи Flash ; однако наши мыши Flash имеют смешанный генетический фон; это исключает пересадку подбородка на спинку между отдельными животными Flash . Кроме того, из-за высокотравматичного характера процедуры аутотрансплантации из области подбородка мы не смогли получить разрешение IACUC на проведение экспериментов по пересадке подбородка на спинку у одного и того же животного.
Вместо этого мы провели следующие дополнительные эксперименты, чтобы усилить связь между доменно-специфической динамикой цикла волос и передачей сигналов BMP / WNT:
a) Мы провели дополнительные эксперименты по пересадке кожи подбородка и дорсальной WT для доноров SCID, чтобы подтвердить выводы о том, что кожа подбородка имеет более быструю внутреннюю динамику цикла волос. Новые результаты прививки теперь показаны на основном Рисунке 3.
b) Мы выполнили углубленный анализ доменно-специфичных данных РНК-seq, уделяя особое внимание раннему третьему телогену.Этот новый анализ теперь показан на основных рисунках 3C — 3F. По сравнению с дорсальной кожей подбородок и кожа вентральной части демонстрируют заметные различия в экспрессии пути BMP (меньше лигандов BMP и больше антагонистов BMP) и пути WNT (меньше антагонистов WNT). Поскольку RNA-seq проводился на тщательно рассеченной коже, наш анализ дополнительно подтверждает, что наблюдаемые различия в передаче сигналов BMP и WNT присущи этим областям кожи.
c) Мы также исследовали паттерны экспрессии lacZ у мышей-репортеров Axin2-lacZ, WNT и BRE-gal, BMP во время второго телогена (основные рисунки с 4G по 4I и приложение 1 — рисунок 21).В соответствии с данными репортера Flash WNT, мы наблюдаем более быстрое увеличение активности репортера Axin2-lacZ в подбородке и вентральной коже по сравнению с дорсальной кожей. Динамический сигнал lacZ локализуется на дермальных сосочках телогенных волосяных фолликулов, где он функционирует как маркер компетентности телогена (Plikus et al., 2011). Мы видим намного больше lacZ + дермальных сосочков на подбородке и вентральной коже во время раннего второго телогена, на день P36. Для мышей-репортеров BRE-gal мы видим меньше волосяных фолликулов lacZ + (экспрессия области выпуклости) в подбородке и вентральной коже по сравнению с дорсальной кожей (рис. 4I).Эти новые результаты подтверждают наши выводы о том, что телогеновые волосяные фолликулы быстрее приобретают статус передачи сигналов с низким BMP и высоким WNT в подбородке и вентральной коже по сравнению с дорсальной кожей.
d) Мы также сравнили второй телоген кожи подбородка, вентральной и дорсальной частей по гистологии, иммуно-гистохимии и FACS. В соответствии с анализом генной онтологии данных RNA-seq (основной рисунок 3E), мы наблюдаем сайт-специфические различия в распределении кожной жировой ткани и скелетных мышц panniculus carnosus (приложение 1 — рисунок 19), а также макрофагов ( Приложение 1 — рисунок 20).Мы предполагаем, что эти региональные различия в составе кожи могут вносить вклад в региональные различия в активности WNT и BMP, которые лежат в основе более быстрой смены волосков на подбородке и вентральной части волос. По общему признанию, потребуется углубленное последующее исследование для выявления и проверки основных сайт-специфических клеточных источников лигандов и антагонистов WNT и BMP. В то же время наше текущее исследование устанавливает четкую динамику цикла волос на конкретных участках и многообещающие региональные различия в микроанатомии кожи, что требует последующих экспериментов.
5) Предположение о том, что вентральная кожа инициирует круговорот волос, интересно и не полностью проверено экспериментально в этой рукописи.
Мы предполагаем, что, как правило, более быстрый цикл волосяного покрова на подбородке и вентральной части кожи помогает достичь более высокой плотности шерсти, что может помочь с повышенной изоляцией и защитой от механического истирания брюшной полости у животных с низкой походкой, таких как мыши. В этой редакции мы изучили распределение плотности косолапых волос между тремя областями у взрослых 4-месячных мышей.Действительно, мы обнаружили, что волосяные фолликулы на подбородке и вентральном телогене постоянно имели больше косолиц (три или более) по сравнению с волосяными фолликулами на дорсальном телогене (в основном, два или один косолапый волос). Эти новые данные теперь показаны в Приложении 1 — рисунок 23, а соответствующие предположения добавлены в Обсуждение. Мы также признали в Обсуждении, что скорость регенерации волос является только одним из факторов, определяющих густоту шерсти, и что на последний также может влиять скорость выпадения волос ( или экзоген), параметр, который непосредственно не тестировался в этой работе. .
6) Волосяной фолликул, безусловно, является хорошей моделью для начала исследования этих проблем, а сочетание эксперимента и моделирования очень действенно. Может ли это применяться к другим обширным тканям, таким как мышцы, кровь, кости и т. Д.? Может быть, это можно обсудить. Очень интересно, что модель предсказывает своего рода самодостаточность регенеративного процесса, при котором он, подобно вечному двигателю, может потенциально продолжать работу без контроля «высшего порядка» или даже без запуска системным сигналом.Может быть, этот аспект можно выделить.
Мы благодарим рецензентов за это прекрасное предложение. В пересмотренном Обсуждении мы добавили, что кожа с ее многочисленными циклическими волосковыми фолликулами является лишь одним, хотя и ярким примером многих органов, осуществляющих масштабную координацию своей физиологической активности. Как упоминалось выше, электрически связанные сердце и желудок демонстрируют крупномасштабную координацию своей сократительной активности из анатомически определенных областей кардиостимулятора. Другие органы, особенно кишечник, имеют много общих черт с кожей — как и кожа, кишечник содержит множество повторяющихся единиц, богатых стволовыми клетками, вдоль ворсинок.Вполне возможно, что координация регенерации более высокого порядка, аналогичная той, что наблюдается в волосистой коже, происходит между ворсинками кишечника.
В отношении регенерации волос существует несколько уровней координации высшего порядка:
a) Координация внутри популяций волос происходит посредством самоорганизации между отдельными волосяными фолликулами (Plikus et al., 2008 и Plikus et al., 2011). Это может дополнительно модулироваться локальной сигнальной макросредой, включая жировые клетки, иммунные клетки и т. Д.
b) Координация между различными анатомическими областями, такими как подбородок, брюшная и дорсальная части кожи, описанная в данном исследовании.
c) Системная гормональная и нейрональная координация. Действительно, существует большое количество литературы о координации роста волос с помощью гуморальных факторов, включая, помимо прочего, пролактин, половые гормоны и симпатические нервы.
У данного вида обновление меха может регулироваться несколькими из вышеперечисленных механизмов, работающих либо параллельно, либо иерархически, когда один механизм перекрывает другой.В обновленной версии «Обсуждение» мы даем краткое изложение этих идей.
7) Учитывая объем работы, проделанной авторами над рисунком 4, сообщение на удивление мало, кроме описания очень сложных и немного сбивающих с толку динамических шаблонов выражения. У авторов действительно есть несколько генетических моделей, используемых в этом исследовании, которые могут помочь им разобраться в функциях некоторых сигнальных путей при определении различных анатомических доменов. Например, в их моделях K14-Noggin, K14-BMP4, K14Cre-Wnt7b fl / fl, K14-Wnt7a, K5trTA-teto-Dkk1 меняются ли частоты циклических циклов кожи подбородка, изменяется ли продолжительность анагена вентральной кожи? Авторы могли выбрать по крайней мере 1 репрезентативную модель сигнальных путей Wnt и BMP, чтобы сделать это.
Мы согласны с этим. Для этого пересмотра мы перенесли большую часть коррелятивного анализа последовательностей РНК в приложение. Кроме того, мы выполнили более глубокий анализ на ранней второй телогеновой РНК-seq (основные рисунки с 4C по 4F). Этот новый анализ более четко показывает, что подбородок и кожа вентральной области экспрессируют более низкие уровни антагонистов WNT и лигандов BMP и более высокие уровни антагонистов BMP, что согласуется с возможностью того, что эти области менее резистентны. Следуя этой корреляции, мы тщательно исследовали образцы окрашивания lacZ у мышей-репортеров Axin2-lacZ WNT и BRE-gal BMP (основные рисунки с 4G по 4I и приложение 1 — рисунок 21).Эти новые данные подтверждают мнение о том, что как кожа подбородка, так и кожа вентрального телогена приобретают статус передачи сигналов с низким BMP и высоким WNT (т.е. менее рефрактерный статус) быстрее по сравнению с кожей спины. Наконец, следуя предложению рецензентов, мы исследовали паттерны роста волос по всей коже у мутантных мышей Krt14-Wnt7a и Krt14-Bmp4 (основные рисунки 4J и 4K и приложение 1 — рисунок22). В соответствии с предполагаемой ролью пути WNT в качестве активатора цикла роста волос по всей коже, мы наблюдаем потерю доминирования в инициации цикла волос от вентральной части к дорсальной, и вначале в дорсальной коже мышей Krt14-Wnt7a появляются участки инициации эктопического роста волос. второго (Приложение 1 — рис. 22) и третьего анагена (рис. 4J).По аналогии, в соответствии с предполагаемой ролью пути BMP в качестве ингибитора цикла роста волос по всей коже, мы наблюдаем замедленное распространение цикла волос с вентральной части на спинку, формирование границ преждевременного роста волос и в целом асимметричные неровные структуры роста волос в Krt14-Bmp4. мышей в начале третьего анагена (рис. 4K).
8) На рисунке 6H процент ушных фолликулов, переходящих в анаген в этих трансгенных моделях, необходимо определить количественно. Это поможет выяснить, насколько эти сигнальные пути доминируют в поддержании покоя кожи уха.
Количественная оценка фенотипа и дополнительные изображения отдельных образцов кожи ушей мышей WT, Krt14-Noggin и Krt14-Wnt7a были добавлены в новое приложение 1 — рисунок 28. Оба мутанта показывают статистически значимое увеличение количества спонтанно вращающихся волосяных фолликулов уха.
https://doi.org/10.7554/eLife.22772.094
Метод регенерации волос — первый метод роста новых фолликулов
Клетки дермального сосочка, показанные здесь внутри луковицы волосяного фолликула человека, были выращены в лаборатории для образования новых фолликулов.Фото: Клэр Хиггинс / Christiano Lab в CUMC
Колумбийские ученые сделали то, о чем до сих пор заявляли только продавцы змеиного масла: они заставили человеческую кожу прорастать новые волосы из недавно сформированных фолликулов.
Ученые говорят, что разработанный ими метод регенерации волос может помочь не только большому количеству мужчин, которые теряют волосы; он также может стать первым подходящим лечением для женщин с истончением волос и для людей с определенными заболеваниями, вызывающими облысение.
В статье, опубликованной в декабрьском выпуске Proceedings of the National Academy of Sciences , исследователи под руководством генетика и профессора дерматологии из Колумбии Анджела Кристиано сообщили, что они создали человеческий волос, используя силу клеток дермального сосочка, тип взрослого человека. стволовые клетки обнаружены, среди прочего, в основании волосяных фолликулов. Ученые извлекли эти клетки из кожи головы семи лысеющих мужчин, умножили их количество в чашках Петри, а затем ввели полученные кластеры стволовых клеток в небольшие участки кожи человека, которые были привиты к спине мышей.В большинстве случаев, пишут исследователи, человеческие волосы возникли из-за ранее гладкой кожи.
Ученые говорят, что необходимо проделать еще больше работы, прежде чем их метод может быть испытан на людях. Волосы, которые они отрастили до сих пор, не особо крепкие и красивые. У них есть идеи, как это улучшить; они считают, что, изменяя свои методы культивирования клеток, они могут вскоре генерировать клетки дермальных сосочков, которые лучше справляются с построением фолликулов при введении обратно в кожу.
«Этот подход может изменить лечение выпадения волос», — говорит Кристиано, эксперт в области генетики роста волос и здоровья кожи. «Современные лекарства от выпадения волос имеют тенденцию замедлять потерю волосяных фолликулов или потенциально стимулировать рост существующих волос, но они не создают новых волосяных фолликулов».
Не только для мужчин
Сегодня единственный способ наложить волосы на голову человека, где их нет, — это трансплантация волос.Наиболее распространенный тип трансплантации включает хирургическое удаление полоски покрытой волосами плоти, обычно с затылка, и пересадку ее на макушку. Процедура болезненна, требует длительного восстановления и не увеличивает общее количество волос, а просто перемещает их. Лечение также не подходит для многих пациентов. Например, женщины, у которых выпадают волосы, склонны видеть равномерное истончение волос на коже головы.
«Около 90 процентов женщин с выпадением волос не являются сильными кандидатами на операцию по трансплантации волос, потому что им не хватает волос для сбора», — говорит Кристиано.«Многие женщины обращаются за пересадкой волос, и по этой причине им отказывают».
Она и ее коллеги с оптимизмом смотрят на то, что их метод в конечном итоге может быть использован для восстановления волос мужчин и женщин с наиболее распространенной формой выпадения волос, вызванной сокращением волосяных фолликулов, а также людей с заболеваниями, вызывающими выпадение волос, например рубцовая алопеция. Они говорят, что это может даже способствовать созданию более реалистичной кожи, созданной с помощью биомедицинских технологий.
«В заменителях кожи, которые наносят ожоговым пациентам, не хватает одного: у них нет нормальных кожных придатков, таких как волосяные фолликулы и потовые железы», — говорит Колин А.Б. Джахода, биолог стволовых клеток из британского Даремского университета и соавтор статьи. «Эта работа может быть не менее важной с точки зрения возможности создания гораздо более качественного и функционального заменителя кожи».
Функциональная сложность ниши стволовых клеток волосяного фолликула и терапевтическое воздействие на дисфункцию ниши для регенерации волос | Journal of Biomedical Science
Волосы образуют барьер, защищающий кожу от внешних воздействий, а также предохраняющий тело от потери температуры.Человеческие волосы, особенно волосы на голове человека, также имеют важные декоративные функции, которые необходимы для социального общения и хорошего самочувствия. Нежелательное выпадение волос может вызвать психосоциальный стресс у пострадавших людей [1]. Регенерация волос зависит от активации стволовых клеток волосяных фолликулов (HFSC) [2,3,4]. Поскольку волосяной фолликул (HF) является неотъемлемой частью кожи [5], его рост и активность HFSC регулируются различными соседними клетками ниши HFSC в коже [6, 7]. Мы классифицируем составляющие ячейки ниши HFSC на 3 группы в соответствии с их функциями, включая сигнализацию, зондирование и ретрансляцию сообщений.Мы рассмотрим, как активность HFSC регулируется различными сигнальными клетками и как сенсорные и передающие сообщения клетки помогают HF инициировать регенеративную попытку перед лицом местного повреждения и изменений внешней среды. В болезненных состояниях мы обсуждаем, как патологические изменения ниши приводят к нарушению регуляции роста волос. Кроме того, мы обсуждаем, как приток или появление несуществующих клеток в нише HFSC влияет на рост волос и истощает HFSC. Мы также подчеркиваем терапевтические последствия патологии ниши с целью предотвращения выпадения волос и стимулирования роста волос.
Структура волосяного фолликула, цикл волос и HFSC
HF — один из немногих органов, подвергающихся циклической инволюции и регенерации на протяжении всей жизни [5, 6, 8, 9]. Структурно HF представляет собой эпителиальный орган, состоящий из двух основных частей: эпителиального цилиндра, состоящего из кератиноцитов и мезенхимальных клеток дермального сосочка (DP) и кожного влагалища (рис. 1) [5, 10]. Во время цикла роста волос HF проходят фазы анагена (рост), катагена (инволюция) и телогена (покоя), а затем снова входят в анаген (рис.1) [5, 8,9,10,11]. Послеродовой цикл и регенерация HFs зависят от сложного реципрокного эпителиально-мезенхимального взаимодействия [6, 12, 13, 14, 15, 16, 17, 18, 19].
Рис. 1
Структура волосяного фолликула, стволовые клетки волосяного фолликула и волосяной цикл. Покоящиеся HFSC расположены в области выпуклости, а примированные HFSC расположены во вторичном зародыше волос. Они временно активируются в раннем анагене, давая начало потомству, которое растет вниз, образуя нижнюю часть HF. HF циклически прогрессируют через катаген (фаза регресса), телоген (фаза покоя) и анаген (фаза роста).Матричные клетки в волосяной луковице активно размножаются и дифференцируются, чтобы поддерживать продолжающееся удлинение стержня волоса в анагене. В катагене волосяная луковица сжимается, и нижняя часть HF регрессирует через постепенно укороченную эпителиальную нить в HF телогена. В телогене HFSC во вторичном зародыше и выпуклости волос остаются инактивированными
За последние 3 десятилетия был достигнут прогресс в понимании того, как регулируется рост HF, в частности, благодаря открытию HFSC [2,3,4, 20 , 21,22].HFSCs сначала идентифицируются как клетки с медленным циклом, удерживающие метку, локализованные в эпителии выпуклости [2, 22]. Помимо этой популяции относительно покоящихся стволовых клеток, HFs содержат другую популяцию примированных стволовых клеток с более быстрой динамикой активации во вторичном волосяном зачатке HFs телогена [3, 5, 23]. Регенерация HF от телогена к анагену подпитывается скоординированной активацией этих двух популяций клеток: сначала активируются примированные HFSCs во вторичном ростке волос, а затем активируются покоящиеся HFSCs в выпуклости [2,3,4,5].
Сигналы и сигнальные клетки в нише HFSC
По определению, HF сам по себе не требует существования окружающих нишевых клеток, чтобы стать HF [24]. Однако интеграция множества окружающих нишевых клеток наделяет HFSC новыми функциями, особенно их способностью реагировать на изменения локальной, системной и даже внешней среды, чтобы начать регенеративную схему или оставаться в покое. В болезненных состояниях патологическая инфильтрация несуществующих клеток в нише HFSC может привести к нарушению регуляции роста волос.Что составляет микросреду, регулирующую активность HFSC и рост волос? В связи с постоянным прогрессом в исследованиях волос, все больше и больше типов клеток (рис. 2), включая клетки DP, жировую ткань, лимфатические сосуды, нервы и иммунные клетки, определяются как вносящие вклад в нишу HFSC [15, 16, 25 , 26,27,28,29,30,31,32,33,34,35,36], раскрывая сложность и изощренность взаимодействия HFSCs с окружающей средой. Поскольку в нише HFSC могут присутствовать как активирующие, так и ингибирующие сигналы, вероятность активации HFSC — это считывание суммы как активирующих, так и ингибирующих сигналов [37, 38].Двумя основными противодействующими сигналами являются костный морфогенетический белок (BMP) и пути передачи сигналов Wnt / β-catenin [28, 37, 39]. Высокий уровень передачи сигналов BMP поддерживает HFSCs в инактивированном состоянии, в то время как передача сигналов Wnt / β-catenin способствует активации HFSC и поддерживает рост HF [17, 28, 37,38,39,40]. Более того, пути передачи сигналов TGF-β2, Foxp1 и онкостатин М также, как было показано, регулируют цикл волос [16, 29, 37, 41]. Факторы, которые способны изменять баланс передачи сигналов Wnt / β-catenin и BMP, могут модулировать активность HFSC, тем самым подавляя или способствуя вступлению в анаген [6, 16, 29, 38, 42].
Рис. 2
Ниша стволовых клеток волосяного фолликула. Ниша HFSC состоит из различных компонентных клеток, таких как дермальный сосочек, преадипоциты, адипоциты, иммунные клетки и нервы. Системные гормоны также прямо или косвенно регулируют HFSC через нишевые клетки HFSC. В нише HFSC присутствуют как активирующие, так и подавляющие сигналы. Вероятность активации HFSC зависит от суммы всех активирующих и тормозных сигналов
Функциональная категоризация — модули сигнализации, восприятия и передачи сообщений в нише HFSC
В дополнение к клеткам ниши, которые обеспечивают либо активирующие, либо тормозящие сигналы в физиологическом состоянии , нишевые клетки с другими функциями также существуют, позволяя HFSCs ощущать локальные, системные и даже внешние изменения окружающей среды, чтобы корректировать свою активность для удовлетворения местных и организменных потребностей [28, 34, 35, 43,44,45].Поскольку дополнительные клетки с разнообразными функциями могут быть включены по отдельности или в комбинации в нишу HFSC, мы думаем, что нишевые клетки могут быть модулированы, и эти модули могут быть кооптированы для создания ниши (Fig. 3). Это аналог конструкции космического корабля. Комбинация различных функциональных модулей увеличивает функциональность космического корабля. Модули ретрансляции сообщений обеспечивали связь между Аполлоном-11 и космическим центром НАСА. Лунный модуль наделил Аполлон-11 важной функцией по высадке человека на Луну.С этой точки зрения мы разделяем нишевые клетки HFSC на 3 функциональные модульные категории: сигнализация, зондирование и ретрансляция сообщений (рис. 3). Сигнальные клетки напрямую обеспечивают активирующие или ингибирующие сигналы для HFSC через секрецию лиганда или межклеточный контакт. Чувствительные клетки обнаруживают изменения локальных сигналов окружающей среды, а затем прямо или косвенно инструктируют HFSCs оставаться в покое или активироваться. Ячейки ретрансляции сообщений способны передавать удаленные сигналы в нишу HFSC, а затем прямо или косвенно модулировать активность HFSC.Следует отметить, что один тип клеток ниши может выполнять более одной функции. Мы предполагаем, что совместное использование различных функциональных модулей в нише стволовых клеток позволяет животным адаптировать свою регенеративную активность к изменяющейся среде и изменяющимся физиологическим потребностям, тем самым улучшая приспособленность организма в процессе эволюции.
Рис. 3
Функциональная категоризация нишевых клеток HFSC. По функциям нишевых ячеек они делятся на 3 группы: сигнальные модули, сенсорные модули, модули ретрансляции сообщений.Эти функционально разные модули собраны в многофункциональную нишу. Сигнальные модули регулируют активность HFSC посредством межклеточного контакта или паракринной секреции. Сенсорные модули обнаруживают сигналы окружающей среды. Модули ретрансляции сообщений передают сигналы от удаленных клеток / тканей к HFSC. Сенсорные модули и модули передачи сообщений могут напрямую сигнализировать HFSC или косвенно регулировать активность HFSC через сигнальные модули.
Сигналы от дермального сосочка для передачи сигналов и усиления функции обработки тестостерона при андрогенной алопеции.
DP-клетки являются важным сигнальным компонентом внутри Ниша HFSC (рис.2). Эпителиально-мезенхимальное взаимодействие необходимо не только для эмбрионального морфогенеза HF, но и для постнатального цикла волос [12, 13, 36, 46]. На эмбриональной стадии взаимодействие со специализированной мезенхимальной нишей дермального конденсата / сосочка HF стимулирует и инструктирует эпителий к последовательному формированию плакоды, зародыша и колышка [46]. Для постнатального фолликулярного эпителиально-мезенхимального взаимодействия, хотя клетки DP предоставляют сигнальные лиганды, такие как TGF-β2 и FGF-7 [3, 16], для активации HFSC для нового цикла волос, сигналы от эпителиальных клеток также необходимы для правильного входа в анаген. [17].После того, как HFs достигают зрелого анагена, DP ведет себя как инструктивная ниша, регулируя пролиферацию и пространственно упорядоченную дифференцировку транзитных усиливающих предшественников для правильного удлинения стержня волоса и поддержания структуры волосяной луковицы [47]. Продолжающаяся активация передачи сигналов Wnt / β-catenin в DP посредством эпителиально-мезенхимального взаимодействия необходима для прогрессирования анагена [48]. Во время катагена DP также важен для рецессии эпителиального цилиндра посредством контролируемого апоптоза / гибели клеток [12].Помимо физиологического цикла, DP может также инструктировать процесс репарации анагена, чтобы избежать попадания катагена, когда HF повреждаются химио- и лучевой терапией [49, 50].
Андрогены являются важным регулятором роста волос с парадоксальным действием на HF в различных областях тела. Андрогены могут стимулировать трансформацию небольших пушковых HF в большие терминальные HF после полового созревания, таких как волосы на бороде, лобке и подмышечных впадинах [51, 52]. Напротив, в коже черепа людей с генетической предрасположенностью к андрогенетической алопеции или облысению по мужскому типу андрогены подавляют рост волос, что приводит к прогрессирующей миниатюризации HF [53].Гиперандрогения у женщин может привести к гирсутизму с чрезмерным ростом волос по мужскому типу [54]. Эти парадоксальные эффекты андрогенов на рост человеческих волос долгое время оставались загадкой [55, 56]. Андрогены действуют через внутриклеточные рецепторы андрогенов. В HF рецепторы андрогенов в основном экспрессируются DP [57, 58]. Напротив, кератиноциты не экспрессируют рецепторы андрогенов и не демонстрируют зависимую от андрогенных рецепторов сигнальную активацию, что позволяет предположить, что кератиноциты не могут быть первичными отвечающими клетками при HF [59, 60].
Алопеция из-за старения HF характеризуется прогрессирующей атрофией HF с миниатюризацией стержня волоса, удлинением телогена и даже потерей всех HF, что приводит к уменьшению количества волос [61, 62]. У мужчин андрогенетическая алопеция является наиболее частым заболеванием преждевременного старения с сердечной недостаточностью. При генетической предрасположенности у пораженных людей клетки DP в области облысения проявляют более высокую активность 5-альфа-редуктазы типа II, фермента, который обычно сильно экспрессируется в простате [58]. Этот фермент превращает тестостерон в дигидротестостерон за счет 5α-снижения уровня тестостерона [57].Дигидротестостерон — более мощный андроген с более высоким сродством, чем тестостерон [63]. Локальная устойчивая стимуляция DP дигидротестостероном нарушает его функции, что приводит к ухудшению роста волос, сокращению анагена и удлинению телогена [55, 56]. Следовательно, несоответствующее усиление функции, т.е. 5α-снижающая способность обрабатывать тестостерон, этих нишевых сигнальных клеток в структурированном распределении является основной причиной андрогенетической алопеции.
Клетки DP из облысения волосистой части головы пациентов с андрогенной алопецией проявляют признаки старения, такие как потеря репликативного потенциала, изменения размера и формы клеток, уменьшение или потеря характерных маркеров / молекулярной сигнатуры [64, 65].Хотя механизмы еще полностью не выяснены, дигидротестостерон, по-видимому, вызывает преждевременное старение DP из-за постоянной активации рецепторов андрогенов. Облысевшие клетки DP не только теряют способность способствовать пролиферации HFSC, но также продуцируют ингибирующие факторы, которые подавляют HFSC и нарушают пролиферацию кератиноцитов [66,67,68,69]. Напр., Передача сигналов Wnt критична для входа в анаген и его прогрессирования [17, 40]. Dkk1, негативный регулятор передачи сигналов Wnt, сверхэкспрессируется облысением DP клеток [68].Повышенная секреция TGF-β1 из DP в катагене способствует переходу от анагена к катагену [12, 70]. Продукция TGF-β1 активируется при облысении DP и может поставить под угрозу пролиферацию кератиноцитов [69]. Кроме того, облысающие клетки DP также производят более высокие воспалительные цитокины, такие как IL-6 [66, 67, 71]. IL-6 не только ингибирует вход в анаген, но также нарушает нормальное развитие анагена [66, 67, 71]. В качестве ключевого мезенхимального сигнального компонента в нише HFSC целевое восстановление нормальных сигнальных функций DP-клеток может быть важной стратегией лечения алопеции.
В настоящее время наиболее эффективным лечением андрогенетической алопеции является подавление местной продукции дигидротестостерона путем ингибирования 5-α-редуктазы. Финастерид и дутастерид — ингибиторы 5α-редуктазы с разной специфичностью и эффективностью [72, 73]. Финастерид в основном ингибирует 5α-редуктазу II типа, основной подтип 5α-редуктазы при HF, тогда как дутастерид подавляет 5α-редуктазы как I, так и II типа. Длительное лечение финастеридом или дутастеридом способствует росту волос у пациентов с андрогенетической алопецией [74, 75].Другой одобренный FDA препарат для лечения облысения — миноксидил [76, 77]. Миноксидил — это средство, открывающее калиевые каналы, первоначально разработанное для лечения гипертонии [78]. Хотя механизмы до сих пор неясны, предполагается, что он способствует росту волос за счет своего воздействия на кровеносные сосуды или калиевые каналы [79].
Тучные клетки, регуляторные Т-клетки, дендритные эпидермальные Т-клетки
HF поддерживает свою собственную отличительную иммунную систему, а взаимодействие HF с иммунными клетками обеспечивает правильный рост волос и защиту от аутоиммунитета [30, 35, 80, 81].Иммунные клетки, включая макрофаги, тучные клетки и Т-клетки, модулируют активность HFSC (рис. 2) [30,31,32, 35]. Тучные клетки обнаруживаются в перифолликулярном компартменте HF [80]. В то время как роль тучных клеток в активации и дифференцировке HFSC все еще неясна, гистохимический и ультраструктурный анализ кожи мышей показал высокий уровень дегрануляции во время перехода от позднего телогена к раннему анагену и от позднего анагена к переходу от раннего катагена [32, 82]. Некоторые молекулы, секретируемые тучными клетками, могут вносить вклад в оборот HF, включая гистамин и серотонин, которые способствуют пролиферации эпидермальных кератиноцитов in situ [83].Также предполагается, что активность тучных клеток вносит свой вклад в нарушения облысения, такие как андрогенная алопеция и рубцовая алопеция [84,85,86].
Было показано, что регуляторные Т-клетки (Treg) находятся в эпителии HFSC и находятся в тесном контакте с HFSC [31]. Treg могут увеличивать пролиферацию и дифференцировку HFSC после травмы выщипывания волос посредством Jagged1 (Jag1, 31). Роль клеток Лангерганса и дендритных эпидермальных Т-клеток (DETC; γδ Т-клетки), которые являются резидентными антигенпредставляющими клетками и Т-клетками соответственно. в модуляции активности HFSC менее определены [80].Клетки Лангерганса и DETC находятся во внешней корневой оболочке HFs [80]. Сообщалось о роли DETC в HFSC в контексте заживления ран. Активированные DETC не только стимулируют пролиферацию эпидермальных стволовых клеток для ускорения заживления ран [87], но также благоприятно способствуют активации HFSC для возобновления роста волос [88].
Макрофаги для передачи сигналов и для восприятия повреждений и силы
Физиологически кластеры резидентных в коже макрофагов могут быть обнаружены в перифолликулярном компартменте и участвуют в регуляции цикла волос [30, 89].Количество резидентных в коже макрофагов CD11b + F4 / 80 + Gr1 — уменьшается из-за апоптоза до наступления анагена [30]. При апоптозе они высвобождают стимулирующие факторы, такие как Wnt7b и Wnt10a, которые способствуют активации и дифференцировке HFSC [30]. Совсем недавно было сообщено, что другая подгруппа TREM2 + дермальных макрофагов (трихофагов) оказывает ингибирующее действие на рост волос [29]. Это исследование было основано на открытии того, что ингибирование передачи сигналов JAK-STAT способствует росту волос за счет нарушения поддержания покоя HFSC [90].Механически онкостатин M действует выше передачи сигналов JAK-STAT5, поддерживая покой HFSC, а онкостатин M продуцируется макрофагами TREM2 + [29]. Истощение этого специфического подмножества макрофагов приводит к преждевременному вступлению в анаген [29].
Макрофаги также выполняют другие функции, включая распознавание повреждений кожи и механической силы. Ранение способствует преждевременному вступлению в анаген кожи [44, 45]. Когда кожа повреждена, макрофаги рекрутируются и активируются через киназу 1, регулирующую сигнал апоптоза (ASK1) [45].Повреждение HF при выщипывании волос также является мощным стимулом для HFSC. Поврежденные выщипыванием волос, HFs рекрутируют макрофаги посредством высвобождения CCL2 [35]. TNF-α, высвобождаемый активированными макрофагами, активирует HFSC, индуцируя AKT-зависимое накопление β-катенина [91]. Следовательно, макрофаги здесь способствуют нише HFSC, чтобы ощущать повреждения HF или повреждения окружающей кожи, чтобы предпринять регенеративную попытку защиты кожи. Кроме того, макрофаги также опосредуют восприятие механических сигналов.Растяжение кожи может поляризовать макрофаги в сторону фенотипа M2 [92]. Прорегенеративные макрофаги M2 стимулируют регенерацию волос за счет паракринной секреции IGF и HGF. Это демонстрирует механическую ось макрофагов в регуляции регенерации тканей. Поскольку в коже существует несколько популяций макрофагов, каждая из которых играет определенную роль в модуляции активации и дифференцировки HFSC, нацеливание на макрофаги может стать будущим направлением для лечения выпадения волос.
Приток аутореагирующих Т-клеток в нишу HFSC нарушает рост волос в очаговой алопеции и плоском лишайнике.
Правильный цикл HF сильно зависит от гомеостаза в поддержании HFSC, а также от неповрежденной иммунной привилегии [81].Коллапс иммунных привилегий в результате факторов окружающей среды или генетической предрасположенности повышает риск иммунной / воспалительной атаки на HF [81, 93, 94]. Активный иммунный ответ с секрецией воспалительных цитокинов, таких как интерферон-γ и TNF-α, безусловно, может нарушить надлежащее поддержание HFSC, что приведет к алопеции [94, 95]. Эти цитокины в большом количестве секретируются лимфоцитами, которые обычно не присутствуют в физиологическом состоянии, включая CD4 и CD8 Т-клетки (αβ Т-клетки), которые окружают или инфильтрируют HF [94, 96].
Одним из наиболее распространенных случаев облысения, опосредованного иммунитетом, является очаговая алопеция. Очаговая алопеция — это аутоиммунная форма облысения, которая может проявляться на коже головы или прогрессировать до полного облысения [97]. Гнездная алопеция обратима, что указывает на то, что HFSC не теряются во время аутоиммунных / воспалительных атак [97]. Однако точный этиопатогенез очаговой алопеции полностью не выяснен. Развитие очаговой алопеции связано с коллапсом иммунной привилегии HF, что впоследствии увеличивает презентацию антигена для исследования Т-клеток, которые распознают эпителиальные HF и / или ассоциированные с меланоцитами антигены как чужеродные и вызывают аутоиммунные ответы против HF [93, 97, 98, 99 100].Продолжаются исследования, пытающиеся определить точный антиген HF и антиген-специфические Т-клетки, участвующие в возникновении очаговой алопеции [99, 101]. Аутоиммунная атака не убивает специфически HFSC, а вместо этого убивает нижнюю переходную часть HF в анагене [97]. Поскольку HFSC сохраняются, удаление этих патогенных Т-клеток из ниши HFSC восстанавливает рост волос. Из-за большого разнообразия клинических проявлений, таких как количество и степень поражения, возраст начала, продолжительность персистирования заболевания и непредсказуемые реакции на лечение, до сих пор отсутствуют универсальные рекомендации по лечению очаговой алопеции.Местные или внутриочаговые стероиды предпочтительны для пациентов с ограниченными заболеваниями, а местный миноксидил может использоваться в качестве адъювантной терапии [102, 103, 104]. У пациентов с обширным выпадением волос можно назначить системные стероиды и другие иммунодепрессанты, такие как метотрексат. Кроме того, иммунотерапия с многократным местным нанесением контактных сенсибилизаторов, таких как дифенилциклопропенон (DPCP), также применялась у пациентов с обширным выпадением волос [102, 105, 106].
Было высказано предположение, что нарушение регуляции Tregs связано с коллапсом иммунной привилегии HF при очаговой алопеции [31, 107, 108].Дефект или отсутствие Treg может привести к неконтролируемой аутоиммунной атаке на клетки HF [108]. Сообщалось также об улучшении очаговой алопеции (возобновление роста волос и уменьшение Т-лимфоцитов CD4 и CD8) при лечении пациентов низкими дозами IL2, чтобы способствовать привлечению Treg в кожу [109].
Недавно было показано, что популяция CD8 + / NKG2D + Т-клеток необходима и достаточна для развития очаговой алопеции [107, 110, 111]. Ответ IFN-γ и несколько цитокинов γ-цепи (γc) значительно усиливаются в коже очаговой алопеции, что может активировать цитотоксическую CD8 + / NKG2D + инфильтрацию Т-клеток.Использование антитела против INF-γ может эффективно блокировать инфильтрацию Т-клеток CD8 + / NKG2D + и предотвращать развитие очаговой алопеции у мышей [110]. В ходе этого исследования авторы идентифицировали низкомолекулярный ингибитор, который может эффективно блокировать передачу сигналов JAK-STAT, важную для функции Т-клеток CD8 + / NKG2D + и обратной очаговой алопеции как у мышей, так и у пациентов [110]. Это исследование впоследствии привело к двум успешным клиническим испытаниям, в которых одобренные FDA ингибиторы JAK, руксолитиниб (специфичный для JAK1 / 2) и тофацитиниб (пан-JAK), использовались для лечения очаговой алопеции от умеренной до тяжелой [112, 113].В связи с этим было начато больше клинических испытаний с целью оптимизации лечения очаговой алопеции с помощью JAK-специфических ингибиторов и различных способов введения [114]. Как описано выше, ингибирование передачи сигналов JAK-STAT может иметь прямое влияние на цикл волос. В то время как мышей, пораженных очаговой алопецией, лечили, было замечено, что у мышей отрастали более густые волосы. Когда лечение применялось к мышам дикого типа на телогене, мыши быстрее входили в анаген и становились более густыми и темными [90]. Эти наблюдения указывают на двойную роль ингибиторов JAK в очаговой алопеции, ингибируя CD8 + / NKG2D + Т-клеток и способствуя пролиферации или дифференцировке HFSC.
Выпадение волос при плоском лишае, также являющееся хроническим воспалительным заболеванием HF, необратимо с окончательным рубцеванием [94, 115]. Lichen planopilaris обычно имеет медленно прогрессирующее течение, проявляющееся отдельными или множественными пятнами перифолликулярной эритемы, шелушением, фолликулярным гиперкератозом и возможной потерей HF [116]. В отличие от очаговой алопеции, при которой цитотоксические Т-клетки нацелены на волосяную луковицу, плоский лишай характеризуется Th2-смещенной инфильтрацией цитотоксических Т-клеток вокруг области выпуклости, в которой находятся HFSC.Предполагается, что избирательный коллапс иммунных привилегий в нише HFSC, возможно, запускаемый интерфероном-γ, вносит вклад в патогенез плоского лишая [94]. Хроническое воспаление ниши может истощать HFSCs, напрямую индуцируя апоптоз HFSC или косвенно изменяя среду ниши до состояния, неблагоприятного для поддержания HFSCs [94, 117]. Истощение HFSC приводит к потере целых фолликулярных структур. С терапевтической точки зрения также нет единого мнения о лечении этого заболевания.Современное лечение в основном основывается на иммунодепрессантах, таких как местные, внутриочаговые или системные стероиды, гидроксихлорохин, циклоспорин и микофенолятмофетил [116, 118]. Предотвращение коллапса иммунной привилегии ниши HFSC может стать будущим направлением лечения и профилактики этого заболевания [94].
Сигналы от жировой ткани и пищевого зондирования
Кожная белая жировая ткань — это высокодинамичная ткань в коже, толщина которой колеблется во время цикла волос [27, 119, 120, 121].Белая жировая ткань дермы утолщается от телогена к анагену, а затем уменьшается в толщине от анагена к переходу от катагена [120]. Увеличение толщины белой жировой ткани дермы во время перехода телогена в анаген в основном обусловлено пролиферацией и дифференцировкой преадипоцитов и гипертрофией созревших адипоцитов [27, 122]. Созревание преадипоцитов с усилением адипогенеза зависит от эпидермальной передачи сигналов Wnt / β-catenin и sonic hedgehog (SHH) [123, 124].Эпидермальная передача сигналов Wnt / β-catenin является инициатором сигнального каскада, который необходим для дифференцировки дермальных адипоцитов [123]. После начала анагена повышенная продукция SHH клетками, усиливающими транзит HF, способствует адипогенезу в преадипоцитах через рецептор γ, активируемый пролифератором пероксисом [124]. Как толщина кожной белой жировой ткани уменьшается во время перехода от анагена к катагену, до сих пор неясно. Поскольку апоптоз зрелых адипоцитов не обнаруживается [27], возможно, что адипоциты могут подвергаться дедифференцировке посредством липолитического или аутофагического процесса [121].Во время перехода от анагена к катагену HF экспрессируют более высокий уровень TGF-β1, который подавляет пролиферацию и увеличивает апоптоз HF [125]. HFs могут индуцировать дедифференцировку адипоцитов посредством передачи сигналов TGF-β1 в катагене [126].
Было показано, что жировая ткань обладает неметаболическими функциями [127]. В костном мозге кроветворение и активность гемопоэтических стволовых клеток подавляются, когда присутствуют более зрелые адипоциты [128, 129]. В коже адипоциты и преадипоциты играют противоположную роль в регуляции активности HFSC во время физиологического цикла волос (рис.2) [127]. Сразу после активации HFSC для инициации роста анагена зрелые адипоциты высвобождают белки BMP для подавления активности HFSC [28]. Это может предотвратить сверхактивацию HFSCs за счет консолидации покоя. С другой стороны, во время перехода от телогена к анагену преадипоциты стимулируют HFSC через паракринную секрецию PDGF [27]. Взаимная передача сигналов и тесное взаимодействие между HFs и жировой тканью подчеркивают взаимозависимость между HFSCs и его нишевыми клетками для поддержания соответствующей тканевой динамики в коже.
Помимо пассивного накопления жира, жировая ткань также выполняет другие неметаболические функции [127]. Во время бактериальной инвазии кожные адипоциты подвергаются реактивному адипогенезу, чтобы увеличить продукцию антимикробных пептидов против бактерий [130]. Мы предполагаем, что адипоциты в нише HFSC могут играть роль в ощущении изменений окружающей среды, таких как системные состояния питания или локальное повреждение кожи. На рост волос влияют системные состояния питания [131]. У человека нарушение роста волос наблюдается у людей с белково-энергетической недостаточностью [132].При экспериментальном ограничении калорийности HFs могут быть задержаны в пролонгированном телогене [5, 133]. Как HFSC обнаруживают системные состояния питания, неясно. Одна из возможностей заключается в том, что HFSC могут непосредственно определять системные изменения в питании. Передача сигналов mTOR является ключевым путем метаболического ответа на состояние питания [134] , и усиленная передача сигналов mTOR важна для активации HFSC в раннем анагене и регенерации после повреждения ионизирующим излучением [135, 136]. HFSCs могут настраивать передачу сигналов mTOR в соответствии с изменениями системного питания.Другая возможность заключается в том, что состояние питания определяется клетками ниши, такими как адипоциты. В кишечнике ограничение калорий снижает активность mTOR в нишевых клетках Панета [137]. Впоследствии клетки Панета сигнализируют кишечным СК об увеличении количества кишечных СК. Клинические наблюдения показывают, что ожирение может отрицательно влиять на рост волос [138]. Адипоциты могут регулировать HFSC через высвобождение адипокинов в соответствии с системным состоянием питания [127].
Сигналы от сенсорных нервов и функция передачи сообщений симпатических нервов для активации HFSC через симпатический контур ipRGC-SCN
HF — это сенсорный орган с высокой иннервируемой способностью.Неинкапсулированные окончания сенсорных нервов окружают HF для механосенсорной функции [139, 140]. В HFs сенсорные нервы иннервируют верхнюю выпуклость, чтобы сформировать сенсорную пилоневральную нишу (Fig. 2) [139, 140]. Посредством секретирования лигандов SHH эта сенсорная пилоневральная ниша поддерживает более высокую сигнальную активность hedgehog в HFSCs верхней области выпячивания [26]. Хотя эта сенсорная пилоневральная ниша не оказывает значительного влияния на регенерацию волос, способность верхних клеток выпячивания восстанавливать повреждение эпидермиса зависит от устойчивой активации передачи сигналов hedgehog [26].
Функция пилоэрекции HFs основана на упорядоченной интеграции симпатических нервов и мышцы arrector pili вокруг HF. Симпатические нервы не только плотно окружают мышцу arrector pili, но также петляют вокруг HFSCs [34, 140]. Интересно, играют ли симпатические нервы вокруг HF двойную роль как в пилоэрекции (мурашки по коже), так и в регуляции HFSC. В костном мозге симпатические нервы контролируют множество функций гемопоэтических SCs, включая их мобилизацию, поддержание молодой функциональной сигнатуры и регенерацию после химиотерапевтического повреждения [141, 142, 143].Симпатические нервы также передают центральные циркадные ритмы в гемопоэтические SCs для их ежедневного ритмичного колебательного выхода из костного мозга [143,144,145]. Клинические наблюдения показали, что гипертрихоз в форме «гемитрихоза» может быть результатом гиперактивности симпатических нервов из-за хирургической травмы грудной клетки [145], что свидетельствует о стимулирующем влиянии симпатических нервов на рост волос. Ранние эксперименты показали, что симпатические нервы могут способствовать прогрессированию анагена после активации HFSCs в физиологическом состоянии [140].Мы обнаружили, что свет может стимулировать рост волос не только напрямую через кожное облучение, но и косвенно через глаза [34, 43, 146]. Световое облучение глаз мышей, сигнал опасности для ночных животных, обнаруживается нетрадиционным фоторецептором меланопсином внутренних светочувствительных ганглиозных клеток сетчатки (ipRGC) (рис. 4) [34]. Световые сигналы передаются через ipRGC в супрахиазматическое ядро для активации системной симпатической системы. Высокий симпатический тонус увеличивает местное высвобождение норадреналина, что впоследствии активирует передачу сигналов hedgehog в HFSC, способствуя их активации.Следовательно, симпатические нервы являются нишевыми воротами для внутренних HFSCs, чтобы общаться с внешним миром посредством ретрансляции внешних световых сигналов в нишу HFSC. С терапевтической точки зрения стимуляция адренергических рецепторов HFSC может способствовать росту волос.
